

IPC分类号 : C12N15/09,C07K7/08,C07K16/00,C07K17/00,G01N30/88,G01N33/53,G01N33/68









专利摘要
肽、使用该肽的人IgA的分析或纯化方法,所述肽的特征在于,含有(X1-3)-C-(X8-10)-C-(X1-3)(式中,X各自独立地为半胱氨酸之外的任意氨基酸残基,C为半胱氨酸残基)所示的由12~18个氨基酸残基组成的氨基酸序列,且可以与人IgA结合。
权利要求
1.一种肽,其特征在于,含有下述式I所示的由12~18个氨基酸残基组成的氨基酸序列,且可以与人IgA结合:
(X1-3)-C-(X8-10)-C-(X1-3) (I)
式中,X各自独立地为半胱氨酸之外的任意氨基酸残基,C为半胱氨酸残基。
2.权利要求1所示的肽,其特征在于,含有下述式II所示的由16~18个氨基酸残基组成的氨基酸序列,且可以与人IgA结合:
(X3)-C-L-(X7-9)-C-(X3) (II)
式中,X各自独立地为半胱氨酸之外的任意氨基酸残基,C为半胱氨酸残基,L为亮氨酸残基,其中,为18个氨基酸残基时,从N末端起第9个和第10个氨基酸残基X独立地为半胱氨酸之外的任意氨基酸残基,或者其任一者或两者缺失。
3.权利要求2所述的肽,其特征在于,含有下述式III所示的由16~18个氨基酸残基组成的氨基酸序列,且可以与人IgA结合:
(X3)-C-L-X-Y-(X1-3)-G-(X2)-V-C-(X3) (III)
式中,X各自独立地为半胱氨酸之外的任意氨基酸残基,C为半胱氨酸残基,L为亮氨酸残基,Y为酪氨酸残基,G为甘氨酸残基,V为缬氨酸残基,其中,为18个氨基酸残基时,从N末端起第9个和第10个氨基酸残基X独立地为半胱氨酸之外的任意氨基酸残基,或者其任一者或两者缺失。
4.权利要求3所述的肽,其特征在于,含有下述式IV所示的由16~18个氨基酸残基组成的氨基酸序列,且可以与人IgA结合:
(X3)-C-L-X-Y-(X1-3)-G-(X2)-V-C-(X3) (IV)
式中,X各自独立地为半胱氨酸之外的任意氨基酸残基,C为半胱氨酸残基,L为亮氨酸残基,Y为酪氨酸残基,G为甘氨酸残基,V为缬氨酸残基,其中,为18个氨基酸残基时,从N末端起第9个和第10个氨基酸残基X独立地为半胱氨酸之外的任意氨基酸残基,或者或者任一者或两者缺失,以及从N末端起第16个和第18个氨基酸残基独立地为疏水性氨基酸残基。
5.权利要求1~4中任一项所述的肽,其中,为18个氨基酸残基时,从N末端起第1~3、5~14、16~18个的各氨基酸残基为:
第1个氨基酸残基= Q, H, K, R, S或P;
第2个氨基酸残基= M, K, R, L, V, A或D;
第3个氨基酸残基= R, L, M或V;
第5个氨基酸残基= L;
第6个氨基酸残基= S, H, Q, T, K, R, N或A;
第7个氨基酸残基= Y;
第8个氨基酸残基= K或R;
第9个氨基酸残基= C之外的任意氨基酸残基、或者缺失;
第10个氨基酸残基= C之外的任意氨基酸残基、或者缺失;
第11个氨基酸残基= G;
第12个氨基酸残基= R, S, T或K;
第13个氨基酸残基= R, M, K, E, N或P;
第14个氨基酸残基= V;
第16个氨基酸残基= L, F, V或I;
第17个氨基酸残基= W, L, R, E, T, S, Q, P或A;或者
第18个氨基酸残基= L, I, Y, A或V。
6.权利要求1~5中任一项所述的肽,其中,上述式I~式IV的肽中,为18个氨基酸残基时,从N末端起第16个氨基酸残基X为亮氨酸或苯丙氨酸残基。
7.权利要求1~6中任一项所述的肽,其中,上述式I~式IV的肽中,为18个氨基酸残基时,从N末端起第18个氨基酸残基X为亮氨酸残基。
8.权利要求1~7中任一项所述的肽,其由以下1)~26)中任一氨基酸序列组成:
1) HMRCLHYKGRRVCFLL(序列编号1)
2) QMRCLSYKGRRVCLWL(序列编号2)
3) HKRCLHYRGRMVCFLI(序列编号3)
4) KRLCLQYKGSKVCFRL(序列编号4)
5) RMRCLTYRGRRVCLEL(序列编号5)
6) SMRCLQYRGSRVCLTL(序列编号6)
7) QKRCLKYKGSRVCFFL(序列编号7)
8) HLRCLRYKGTRVCFSL(序列编号8)
9) HVRCLSYKGREVCVQL(序列编号9)
10) PRMCLHYKGRRVCIPY(序列编号10)
11) HVRCLRYRGKNVCFLL(序列编号11)
12) SDVCLRYRGRPVCFQV(序列编号15)
13) RDVCLRYRGRPVCFQV(序列编号16)
14) HDVCLRYRGRPVCFQV(序列编号17)
15) SMVCLRYRGRPVCFQV(序列编号19)
16) SAVCLRYRGRPVCFQV(序列编号20)
17) SDVCLNYRGRPVCFQV(序列编号24)
18) SDVCLHYRGRPVCFQV(序列编号25)
19) SDVCLAYRGRPVCFQV(序列编号26)
20) SDVCLRYRGRPVCFRV(序列编号37)
21) SDVCLRYRGRPVCFLV(序列编号38)
22) SDVCLRYRGRPVCFAV(序列编号39)
23) SDVCLRYRGRPVCFQL(序列编号41)
24) SDVCLRYRGRPVCFQA(序列编号42)
25) HMVCLAYRGRPVCFAL(序列编号43)
26) HMVCLSYRGRPVCFSL(序列编号44)。
9.权利要求1~8中任一项所述的肽,其中,肽在2个半胱氨酸(C)残基间形成有二硫键。
10.权利要求1~9中任一项所述的肽,其与IgA的血清型(单体)和分泌型(二聚体)结合。
11.权利要求1~10中任一项所述的肽,其结合有标记标记。
12.融合蛋白质,其由权利要求1~11中任一项所述的肽与所连接的蛋白质组成。
13.固定化肽,其是将权利要求1~11中任一项所述的肽与固相结合而成。
14.核酸,其编码权利要求1~11中任一项所述的肽。
15.IgA的纯化方法,其包括使权利要求1~11中任一项所述的肽或权利要求13所述的固定化肽与IgA结合,以及使结合的IgA游离而回收IgA。
16.IgA的检测方法,其包括使样品中的IgA与权利要求1~11中任一项所述的肽或权利要求13所述的固定化肽结合而检测结合的IgA。
17.用于人IgA的分析或纯化的试剂盒,其含有权利要求1~11中任一项所述的肽或权利要求13所述的固定化肽的至少1种。
18.IgA分离用柱,其含有权利要求13所述的固定化肽。
说明书
技术领域
本发明涉及由随机肽文库得到的人IgA结合性肽以及利用该肽的IgA的分析方法或纯化方法。
背景技术
免疫球蛋白A(IgA)不仅在粘膜免疫中是重要的抗体,而且在血中是继免疫球蛋白G(IgG)的第2个主要的抗体家族,发挥对于细菌、病毒感染的防御功能。IgA中存在具有二聚体结构的分泌型IgA(sIgA)与单体结构的IgA(mIgA)。sIgA具有通过SS键所形成的连接链(J-链)连接的二聚体结构,分泌至粘液中,但mIgA多数存在于血液中。另外,IgA中存在主要是绞链区的长度不同的2种亚型(IgA1与IgA2),IgA2缺少富含Pro的13个残基的区域。对于IgA在医药品中的功能,由于在感染免疫中的重要性,在粘膜疫苗的开发中受到关注(非专利文献1和2),由于血液中的IgA特别是被报道有嗜中性粒细胞介导的对于癌细胞的ADCC(非专利文献3和4),因此,与作为癌症或自身免疫疾病的治疗药的临床应用不断扩大的抗体药物形式的IgG相同地,IgA也被期待作为靶向癌症的抗体药物(非专利文献5)。
但是,作为妨碍IgA开发为医药品的主要原因之一,可举出无法确立如IgG制造中的蛋白质A/G亲和柱那样能够对应于工业性、制药规模的纯化方法。以往,作为IgA的纯化方法,报道有数种方法(非专利文献6)。例如,报道了利用识别IgA1特异性糖链的外源凝集素Jackalin(非专利文献7)或作为蛋白质A模拟合成配体的TG19318(非专利文献8)来纯化IgA的方法,但它们由于结合能力或特异性中的问题,因而在利用上存在限制。另外,从链球菌来源的表面蛋白质M 蛋白质家族(非专利文献9)的成员中发现了IgA结合蛋白质(非专利文献10、11和专利文献1),但与IgG等血清中的其它蛋白质的相互作用等(非专利文献12)成为问题,因而作为IgA特异性亲和性配体使用时存在障碍。另一方面,Sandin等人取出由该Streptococcal Sir22 (M22)蛋白中的48个残基组成的功能区肽(Streptococcal IgA-结合肽, Sap),利用经由Cys的SS键进行二聚体化,由此报道了与原来的Sir22蛋白质相比亲和性(以Kd值计为3~4nM)降低,但亲和性较高(以Kd值计为20nM)的IgA的纯化用亲和配体(非专利文献13、专利文献2)。实际上,该配体与IgA的Fc结合,可以用于sIgA、mIgA两者的纯化、以及抗原特异性的IgA1、IgA2单克隆抗体的检测。
与上述IgA结合蛋白质相同地,对于IgG,本发明人开发了IgG结合性肽(专利文献3)。
现有技术文献
专利文献
专利文献1:国际公开第WO 1992/017588
专利文献2:国际公开第WO 2000/063383
专利文献3:国际公开第WO 2008/054030
非专利文献
非专利文献1:Holmgren, J. (1991) Fems Microbiology Immunology 89(1), 1-9
非专利文献2:Holmgren, J., and Czerkinsky, C. (2005) Nat. Med. 11(4), S45-S53
非专利文献3:Dechant, M., Beyer, T., Schneider-Merck, T., Weisner, W., Peipp, M., vande Winkel, J. G., and Valerius, T. (2007) J Immunol 179(5), 2936-2943
非专利文献4:Zhao, J., Kuroki, M., Shibaguchi, H., Wang, L., Huo, Q., Takami, N., Tanaka, T., Kinugasa, T., and Kuroki, M. (2008) Oncol. Res. 17(5), 217-222
非专利文献5:Beyer, T., Lohse, S., Berger, S., Peipp, M., Valerius, T., and Dechant, M. (2009) Journal of Immunological Methods 346(1-2), 26-37
非专利文献6:Pack, T. D. (2001) Current protocols in Immunology / edited by John E. Coligan et al Chapter 2, Unit 2 10B
非专利文献7:Kondoh, H., Kobayashi, K., and Hagiwara, K. (1987) Molecular immunology 24(11), 1219-1222
非专利文献8:Palombo, G., De Falco, S., Tortora, M., Cassani, G., and Fassina, G. (1998) J Mol Recognit 11(1-6), 243-246
非专利文献9:Frithz, E., Heden, L. O., and Lindahl, G. (1989) Molecular Microbiology 3(8), 1111-1119
非专利文献10:Russell-Jones, G. J., Gotschlich, E. C., and Blake, M. S. (1984) The Journal of Experimental Medicine 160(5), 1467-1475
非专利文献11:Lindahl, G., Akerstrom, B., Vaerman, J. P., and Stenberg, L. (1990) European Journal of Immunology 20(10), 2241-2247
非专利文献12:Stenberg, L., O'Toole, P. W., Mestecky, J., and Lindahl, G. (1994) TheJournal of Biological Chemistry 269(18), 13458-13464
非专利文献13:Sandin, C., Linse, S., Areschoug, T., Woof, J. M., Reinholdt, J., and Lindahl, G. (2002) J Immunol 169(3), 1357-1364。
发明内容
发明要解决的技术问题
本发明的目的在于提供对人IgA特异性或选择性地具有结合性的肽。
另外,本发明的目的还在于提供使用该肽来纯化或分析(例如,检测或定量)人IgA的方法。
用于解决技术问题的手段
人IgA如背景技术所述存在于粘膜和血中,在对于感染等的防御中发挥重要的功能。由于这种特性,IgA被作为抗体药物用于感染疾病、肿瘤等疾病的治疗中。本发明鉴于上述情况而完成,通过提供对可以特异性或选择性地与人IgA结合的肽,认为可用于能过作为药物使用的IgA的纯化和分析。
对本发明进行归纳,则具有以下的特征。
[1] 一种肽,其特征在于,含有下述式I:
(X1-3)-C-(X8-10)-C-(X1-3) (I)
(式中,X各自独立地为半胱氨酸之外的任意氨基酸残基,C为半胱氨酸残基)
所示的由12~18个氨基酸残基组成的氨基酸序列,且可以与人IgA结合。
[2] 上述[1]所述的肽,其特征在于,含有下述式II:
(X3)-C-L-(X7-9)-C-(X3) (II)
(式中,X各自独立地为半胱氨酸之外的任意氨基酸残基,C为半胱氨酸残基,L为亮氨酸残基,其中,为18个氨基酸残基时,从N末端起第9个和第10个氨基酸残基X独立地为半胱氨酸之外的任意氨基酸残基,或者其任一者或两者缺失)
所示的由16~18个氨基酸残基组成的氨基酸序列,且可以与人IgA结合。
[3] 上述[2]所述的肽,其特征在于,含有下述式III:
(X3)-C-L-X-Y-(X1-3)-G-(X2)-V-C-(X3) (III)
(式中,X各自独立地为半胱氨酸之外的任意氨基酸残基,C为半胱氨酸残基,L为亮氨酸残基,Y为酪氨酸残基,G为甘氨酸残基,V为缬氨酸残基,其中,为18个氨基酸残基时,从N末端起第9个和第10个氨基酸残基X独立地为半胱氨酸之外的任意氨基酸残基,或者其任一者或两者缺失)
所示的由16~18个氨基酸残基组成的氨基酸序列,且可以与人IgA结合。
[4] 上述[3]所述的肽,其特征在于,含有下述式IV:
(X3)-C-L-X-Y-(X1-3)-G-(X2)-V-C-(X3) (IV)
(式中,X各自独立地为半胱氨酸之外的任意氨基酸残基,C为半胱氨酸残基,L为亮氨酸残基,Y为酪氨酸残基,G为甘氨酸残基,V为缬氨酸残基,其中,为18个氨基酸残基时,从N末端起第9个和第10个氨基酸残基X独立地为半胱氨酸之外的任意氨基酸残基,或者或者任一者或两者缺失,以及从N末端起第16个和第18个氨基酸残基独立地为疏水性氨基酸残基)
所示的由16~18个氨基酸残基组成的氨基酸序列,且可以与人IgA结合。
[5] 上述[1]~[4]中任一项所述的肽,其中,为18个氨基酸残基时,从N末端起第1~3、5~14、16~18个的各氨基酸残基为:
第1个氨基酸残基= Q, H, K, R, S或P;
第2个氨基酸残基= M, K, R, L, V, A或D;
第3个氨基酸残基= R, L, M或V;
第5个氨基酸残基= L;
第6个氨基酸残基= S, H, Q, T, K, R, N或A;
第7个氨基酸残基= Y;
第8个氨基酸残基= K或R;
第9个氨基酸残基= C之外的任意氨基酸残基、或者缺失;
第10个氨基酸残基= C之外的任意氨基酸残基、或者缺失;
第11个氨基酸残基= G;
第12个氨基酸残基= R, S, T或K;
第13个氨基酸残基= R, M, K, E, N或P;
第14个氨基酸残基= V;
第16个氨基酸残基= L, F, V或I;
第17个氨基酸残基= W, L, R, E, T, S, Q, P或A;或者
第18个氨基酸残基= L, I, Y, A或V。
[6] 上述[1]~[5]中任一项所述的肽,其中,上述式I~式IV的肽中,为18个氨基酸残基时,从N末端起第16个氨基酸残基X为亮氨酸或苯丙氨酸残基。
[7] 上述[1]~[6]中任一项所述的肽,其中,上述式I~式IV的肽中,为18个氨基酸残基时,从N末端起第18个氨基酸残基X为亮氨酸残基。
[8] 上述[1]~[7]中任一项所述的肽,其中,其由以下1)~26)中任一氨基酸序列组成:
1) HMRCLHYKGRRVCFLL(序列编号1)
2) QMRCLSYKGRRVCLWL(序列编号2)
3) HKRCLHYRGRMVCFLI(序列编号3)
4) KRLCLQYKGSKVCFRL(序列编号4)
5) RMRCLTYRGRRVCLEL(序列编号5)
6) SMRCLQYRGSRVCLTL(序列编号6)
7) QKRCLKYKGSRVCFFL(序列编号7)
8) HLRCLRYKGTRVCFSL(序列编号8)
9) HVRCLSYKGREVCVQL(序列编号9)
10) PRMCLHYKGRRVCIPY(序列编号10)
11) HVRCLRYRGKNVCFLL(序列编号11)
12) SDVCLRYRGRPVCFQV(序列编号15)
13) RDVCLRYRGRPVCFQV(序列编号16)
14) HDVCLRYRGRPVCFQV(序列编号17)
15) SMVCLRYRGRPVCFQV(序列编号19)
16) SAVCLRYRGRPVCFQV(序列编号20)
17) SDVCLNYRGRPVCFQV(序列编号24)
18) SDVCLHYRGRPVCFQV(序列编号25)
19) SDVCLAYRGRPVCFQV(序列编号26)
20) SDVCLRYRGRPVCFRV(序列编号37)
21) SDVCLRYRGRPVCFLV(序列编号38)
22) SDVCLRYRGRPVCFAV(序列编号39)
23) SDVCLRYRGRPVCFQL(序列编号41)
24) SDVCLRYRGRPVCFQA(序列编号42)
25) HMVCLAYRGRPVCFAL(序列编号43)
26) HMVCLSYRGRPVCFSL(序列编号44)。
[9] 上述[1]~[8]中任一项所述的肽,其中,肽在2个半胱氨酸(C)残基间形成有二硫键。
[10] 上述[1]~[9]中任一项所述的肽,其与IgA的血清型(单体)和分泌型(二聚体)结合。
[11] 上述[1]~[10]中任一项所述的肽,其结合有标记。
[12] 融合蛋白质,其由上述[1]~[11]中任一项所述的肽与所连接的蛋白质组成。
[13] 固定化肽,其是将上述[1]~[11]中任一项所述的肽与固相结合而成。
[14] 核酸,其编码上述[1]~[11]中任一项所述的肽。
[15] IgA的纯化方法,其包括使上述[1]~[11]中任一项所述的肽或上述[13]所述的固定化肽与IgA结合,以及使结合的IgA游离而回收IgA。
[16] IgA的检测方法,其包括使样品中的IgA与上述[1]~[11]中任一项所述的肽或上述[13]所述的固定化肽结合而检测结合的IgA。
[17] 用于人IgA的分析或纯化的试剂盒,其含有上述[1]~[11]中任一项所述的肽或上述[13]所述的固定化肽的至少1种。
[18] IgA分离用柱,其含有上述[13]所述的固定化肽。
本说明书包含作为本申请优选权基础的日本专利申请2010-118508号的说明书和/或附图中记载的内容。
本发明的人IgA结合性肽具有下述优点:与IgG、IgM和IgE相比,对IgA具有更高的选择性,可以与人IgA结合。这意味着可以由例如人血清等中选择性地分离IgA。
附图说明
[图1]显示5次淘选后所得噬菌体的利用ELISA的相对于人IgA的结合特异性。
[图2]显示人IgA结合噬菌体克隆的利用ELISA的结合特异性。
[图3]显示hIgA-2 肽模体(motif)的氨基酸序列与DNA序列。
[图4]显示来自部分突变文库的人IgA特异性的噬菌体的浓缩。
[图5]显示hIgA-2合成肽的结合特异性。其中,IgG-BP表示人IgG特异性的肽、LF-A1表示人乳铁传递蛋白特异性肽。
[图6]表示利用表面等离子的hIgA-2合成肽对人IgA的结合分析的结果。
[图7]表示利用固定化了hIgA-2肽的柱的人IgA的特异性吸附和洗脱。
[图8]表示利用固定化了hIgA-2肽的柱从人血清中纯化IgA。
[图9]表示由固定化了hIgA-2肽的柱回收的IgA的SDS-PAGE(左侧)与Westernblotting(右侧)。
[图10]表示用于分离对hIgA的亲和性高的模体的最优化文库的设计。
[图11]表示通过从最优化文库中淘选来浓缩hIgA特异性的噬菌体。
[图12]表示A3-1展示噬菌体克隆的利用ELISA的对hIgA的结合特异性。
[图13]表示利用表面等离子共振分析的A3-1(Opt2)合成肽对人IgA的结合分析的结果。
[图14]表示利用固定化了A3-1(Opt2)合成肽的柱从人血清中纯化IgA。
具体实施方式
本发明中,本发明人发现的对人IgA具有特异性或选择性的结合性的肽是参考通过T7噬菌体展示系统构建的在分子内含有1个二硫键的随机肽文库( Sakamoto, K., Ito, Y., Hatanaka, T., Soni, P. B., Mori, T., and Sugimura, K. (2009) The Journal of biological chemistry 284(15), 9986-9993),利用生物淘选法由新设计、构建的文库分离而得的,此时所得的4种特异性克隆可观察到相互共通的序列的相同性,基于该序列进行各种置换或缺失而制备的合成肽显示对于IgA的优异的特异性与亲和性。对这些肽的结合IgA所必需的残基进行鉴定,使得增强亲和性的方法以及使用该肽从人血清中纯化IgA中的应用成为可能。本发明的IgA结合性肽是最为精简、即小的IgA结合性肽,是12或13个残基,即使与非专利文献13中记载的Streptococcus Sir22 (M22)来源的约50个残基的Sap肽(C末端具有1个Cys)相比,也要小至1/4左右,由此,可期待以低成本的肽为基础来构建IgA的纯化系统。
以下,对本发明进一步详细说明。
具体地,对本发明的IgA结合性肽、利用该肽的IgA的纯化方法和分析方法、用于这种IgA纯化或检测的试剂盒进行说明。
(IgA结合性肽)
本发明的肽是从含有大量随机肽的噬菌体文库之中作为与人IgA特异性或选择性地具有结合性的肽而筛选出的,与以往公知的如非专利文献13中记载的多肽相比,其来源、一次结构不同。
本说明书中使用的人IgA是指IgA1和/或IgA2。
即,对于本发明的肽,作为最为广义的一次结构,是下述肽,该肽的特征在于,含有下述式I:
(X1-3)-C-(X8-10)-C-(X1-3) (I)
(式中,X各自独立地为半胱氨酸之外的任意氨基酸残基,C为半胱氨酸残基)
所示的由12~18个氨基酸残基组成的氨基酸序列,且可以与人IgA结合。
上述式中,N末端或C末端的X1-3的表述意指半胱氨酸(C或Cys)之外的独立的任意氨基酸残基X连续1~3个,构成其的氨基酸残基为相同或不同的残基,但优选由3个均不为相同残基的序列组成。相同地,X8-10也意指半胱氨酸(C或Cys)之外的独立的任意氨基酸残基X连续8~10个,构成其的氨基酸残基为相同或不同的残基,但优选由3个以上不为相同残基的序列组成。另外,以下式中有X3、X7-9、X2等表述,它们也具有同样的含义。
式I的2个半胱氨酸残基可以形成二硫键从而形成环状肽。通常,式I的肽形成有二硫键。
式I的肽的氨基酸序列中,进一步特定了氨基酸残基X的式II、式III和式IV所示的肽示于以下。
即,式II所示的肽的特征在于,含有
(X3)-C-L-(X7-9)-C-(X3) (II)
(式中,X各自独立地为半胱氨酸之外的任意氨基酸残基,C为半胱氨酸残基,L为亮氨酸残基,其中,为18个氨基酸残基时,从N末端起第9个和第10个氨基酸残基X独立地为半胱氨酸之外的任意氨基酸残基,或者其任一者或两者缺失)
所示的由16~18个氨基酸残基组成的氨基酸序列,且可以与人IgA结合。
式III所示的肽的特征在于,含有
(X3)-C-L-X-Y-(X1-3)-G-(X2)-V-C-(X3) (III)
(式中,X各自独立地为半胱氨酸之外的任意氨基酸残基,C为半胱氨酸残基,L为亮氨酸残基,Y为酪氨酸残基,G为甘氨酸残基,V为缬氨酸残基,其中,为18个氨基酸残基时,从N末端起第9个和第10个氨基酸残基X独立地为半胱氨酸之外的任意氨基酸残基,或者其任一者或两者缺失)
所示的由16~18个氨基酸残基组成的氨基酸序列,且可以与人IgA结合。
式IV所示的肽的特征在于,含有
(X3)-C-L-X-Y-(X1-3)-G-(X2)-V-C-(X3) (IV)
(式中,X各自独立地为半胱氨酸之外的任意氨基酸残基,C为半胱氨酸残基,L为亮氨酸残基,Y为酪氨酸残基,G为甘氨酸残基,V为缬氨酸残基,其中,为18个氨基酸残基时,从N末端起第9个和第10个氨基酸残基X独立地为半胱氨酸之外的任意氨基酸残基,或者或者任一者或两者缺失,以及从N末端起第16个和第18个氨基酸残基独立地为疏水性氨基酸残基)
所示的由16~18个氨基酸残基组成的氨基酸序列,且可以与人IgA结合。
上述式II~IV的肽的氨基酸序列中,为18个氨基酸残基时,从N末端起第9个和第10个氨基酸残基X优选均缺失,这样的肽由16个氨基酸长度组成。
本说明书中使用的“为18个氨基酸残基时”是在将肽的氨基酸残基按氨基酸位置编号来称呼时,为了从作为最长氨基酸长度的18个残基的N末端起依次由第1个至第18个进行编号,而方便地表现的用语。
进而,上述各式的肽的氨基酸序列的半胱氨酸(C)之外的氨基酸残基,即,为18个氨基酸残基时,从N末端起第1~3、5~14、16~18个各氨基酸残基优选选自以下。其中,各大写的字母表示氨基酸的单字母符号。
第1个氨基酸残基= Q, H, K, R, S或P、优选H
第2个氨基酸残基= M, K, R, L, V, A或D、优选M
第3个氨基酸残基= R, L, M或V、优选V
第5个氨基酸残基= L
第6个氨基酸残基= S, H, Q, T, K, R, N或A、优选S或A
第7个氨基酸残基= Y
第8个氨基酸残基= K或R、优选R
第9个氨基酸残基= C之外的任意氨基酸残基、或者缺失、优选缺失
第10个氨基酸残基= C之外的任意氨基酸残基、或者缺失、优选缺失
第11个氨基酸残基= G
第12个氨基酸残基= R, S, T或K、优选R
第13个氨基酸残基= R, M, K, E, N或P、优选P
第14个氨基酸残基= V
第16个氨基酸残基= L, F, V或I、优选L或F
第17个氨基酸残基= W, L, R, E, T, S, Q, P或A、优选S或A
第18个氨基酸残基= L, I, Y, A或V、优选L
本发明的肽的数个具体例列举为以下1)~26),但并不受其限定。这种肽相对于人IgA,与其它种系的免疫球蛋白相比,均具有显著更高的结合特异性或结合选择性。
1) HMRCLHYKGRRVCFLL(序列编号1)
2) QMRCLSYKGRRVCLWL(序列编号2)
3) HKRCLHYRGRMVCFLI(序列编号3)
4) KRLCLQYKGSKVCFRL(序列编号4)
5) RMRCLTYRGRRVCLEL(序列编号5)
6) SMRCLQYRGSRVCLTL(序列编号6)
7) QKRCLKYKGSRVCFFL(序列编号7)
8) HLRCLRYKGTRVCFSL(序列编号8)
9) HVRCLSYKGREVCVQL(序列编号9)
10) PRMCLHYKGRRVCIPY(序列编号10)
11) HVRCLRYRGKNVCFLL(序列编号11)
12) SDVCLRYRGRPVCFQV(序列编号15)
13) RDVCLRYRGRPVCFQV(序列编号16)
14) HDVCLRYRGRPVCFQV(序列编号17)
15) SMVCLRYRGRPVCFQV(序列编号19)
16) SAVCLRYRGRPVCFQV(序列编号20)
17) SDVCLNYRGRPVCFQV(序列编号24)
18) SDVCLHYRGRPVCFQV(序列编号25)
19) SDVCLAYRGRPVCFQV(序列编号26)
20) SDVCLRYRGRPVCFRV(序列编号37)
21) SDVCLRYRGRPVCFLV(序列编号38)
22) SDVCLRYRGRPVCFAV(序列编号39)
23) SDVCLRYRGRPVCFQL(序列编号41)
24) SDVCLRYRGRPVCFQA(序列编号42)
25) HMVCLAYRGRPVCFAL(序列编号43)
26) HMVCLSYRGRPVCFSL(序列编号44)。
上述26种肽之中,序列编号43(A3-1(Opt1))和序列编号44(A3-1(Opt2))的肽尤其具有与人IgA(也记载为“hIgA”)的高亲和性。
如前所述,本发明涉及的上述式的肽的特征在于,各氨基酸序列中具有离开的2个半胱氨酸(C)残基,且半胱氨酸残基被配置为能够在该半胱氨酸残基间形成二硫键,优选的肽是2个半胱氨酸残基形成二硫键从而形成环状肽,各半胱氨酸残基的N末端侧和C末端侧具有1~3个、优选3个半胱氨酸之外的任意氨基酸残基。第1~3、16~18个的这类氨基酸残基如上述例示。
本发明的肽与人IgA的结合亲和性与其它人免疫球蛋白(IgG、IgE或IgM)相比,高约10倍以上、优选约50倍以上、更优选约200倍以上。关于本发明的肽与人IgA的结合的离解常数(Kd),可以通过表面等离子共振谱分析(使用例如BIACORE系统(蛋白质相互作用分析))来决定,例如,为1×10-5M~小于1×10-7M、优选为小于1×10-8M、更优选为小于1×10-9M、进一步优选为小于1×10-10M。
使用固定化于固相的本发明的肽,在实际中尝试与人血清中的IgA的结合时,明确了与IgA的血清型(单体)和分泌型(二聚体)结合的事实,暗示了任何一种IgA的分离均可进行。
本发明的肽可以通过惯用的液相合成法、固相合成法等肽合成法、利用自动肽合成机的肽合成等来制造(Kelley et al., Genetics Engineering Principles and Methods, Setlow, J.K. eds., Plenum Press NY. (1990) Vol.12, p.1-19;S tewart et al., Solid-Phase Peptide Synthesis (1989) W.H. Freeman Co.; Houghten, Proc. Natl. Acad. Sci. USA (1985) 82: p.5132,《新生化学実験講座1 蛋白质IV》(1992)日本生化学会编,东京化学同人,东京,日本)。或者,也可以通过使用了编码本发明的肽的核酸的基因重组方法或噬菌体展示法等来制造肽。例如,可以将编码本发明的肽的氨基酸序列的DNA整合至表达载体中,导入宿主细胞中进行培养,由此制造目标肽。 制造的肽可以通过常法,例如,凝胶过滤层析、离子交换柱层析、亲和层析、反相柱层析、HPLC等层析、硫酸铵分级沉淀、超滤、免疫吸附法等进行回收或纯化。
肽合成是准备保护了各氨基酸的、除要结合的α-氨基与α-羧基之外的官能团的氨基酸类,在各氨基酸的α-氨基与α-羧基之间进行肽键形成反应。通常,将位于肽的C末端的氨基酸残基的羧基通过适当的间隔体(スペーサー)或连接体(リンカー)与固相结合。将以上得到的二肽的氨基末端的保护基选择性地除去,在与下一氨基酸的α-羧基之间形成肽键。连续地进行这样的操作而制造侧基被保护的肽,最后除去全部保护基,由固相分离。保护基的种类或保护方法、肽键法的具体内容详细记载于上述文献中。
基因重组方法包括将编码本发明肽的DNA插入适当的表达载体中,将载体導入至适当的宿主细胞中,培养细胞,由细胞内或由细胞外液回收目标肽。载体没有限定,例如,为质粒、噬菌体、粘粒、噬菌粒、病毒等载体。质粒载体没有限定,但可举出来源于大肠杆菌的质粒(例如pET22b(+)、pBR322、pBR325、pUC118、pUC119、pUC18、pUC19、pBluescript等)、来源于枯草杆菌的质粒(例如pUB110、pTP5等)、来源于酵母的质粒(例如YEp13、YCp50等)等。噬菌体载体没有限定,但可举出T7噬菌体展示载体(T7Select10-3b、T7Select1-1b、 T7Select1-2a、T7Select1-2b、T7Select1-2c等(Novagen))、λ噬菌体载体(Charon4A、 Charon21A、EMBL3、EMBL4、λgt10、λgt11、λZAP、λZAPII等)。病毒载体没有限定,可举出例如:反转录病毒、腺病毒、腺相关病毒、牛痘病毒、仙台病毒等动物病毒、杆状病毒等昆虫病毒等。粘粒载体没有限定,可举出Lorist 6、Charomid9-20、Charomid9-42等。噬菌粒载体没有限定,已知例如pSKAN、pBluescript、pBK、pComb3H等。载体中可以包含使目标DNA为可表达的调节序列、或用于筛选含有目标DNA的载体的选择标记物、用于插入目标DNA的多克隆位点等。这类调节序列中含有启动子、增强子、终止子、S-D序列或核糖体结合位点、复制起始区、poly(A)位点等。另外,选择标记物可以使用,例如,氨苄青霉素抗性基因、新霉素抗性基因、卡那霉素抗性基因、二氢叶酸还原酶基因等。用于导入载体的宿主细胞为大肠杆菌、枯草杆菌等细菌、酵母细胞、昆虫细胞、动物细胞(例如,哺乳动物细胞)、植物细胞等,对这些细胞的转化或转染包括例如:磷酸钙法、电穿孔法、脂质体转染法、粒子枪法、PEG法等。培养转化细胞的方法按照宿主生物的培养中使用的通常方法来进行。例如,在大肠杆菌或酵母细胞等微生物的培养中,含有宿主微生物可进行同化的碳源、氮源、无机盐类等。为了使本发明的肽的回收变得容易,优选使表达生成的肽分泌至细胞外。因此,将编码可使肽从该细胞分泌的肽序列的DNA结合于编码目标肽的DNA的5'末端侧。转移至细胞膜的融合肽被信号肽酶切断,从而使目标肽分泌释放至培养基中。或者,也可以回收蓄积于细胞内的目标肽。此时,将细胞物理地或化学地破坏,使用蛋白质纯化技术来回收目标肽。
因此,本发明进一步还涉及编码本发明的肽的核酸。其中,核酸包括DNA或RNA(例如mRNA)。
为了可以检测IgA,本发明的肽还可以被标记。标记没有限定,包括例如:荧光色素、化学发光色素、酶、放射性同位素、荧光蛋白质、生物素等。优选的标记的例子为荧光素、FITC等荧光素衍生物、若丹明、四甲基若丹明等若丹明衍生物、德克萨斯红等荧光色素。
对于本发明的肽,也可以使之与任意蛋白质融合。蛋白质只要是GFP(绿色荧光蛋白质)之类的荧光蛋白质、过氧化物酶等酶等,则可将该蛋白质用作标记。此时,可以根据需要通过适当的连接体通过基因重组方法以融合蛋白质的形式制作本发明的肽和该蛋白质。此时,本发明的肽应该以无损与人IgA的结合性的方式制作融合蛋白质。
本发明的肽还可以进一步以能够用于人IgA的分离纯化、分析等的方式固定化在能够填充于亲和柱的固相上。
适用于将肽固定化的固相没有限定,可举出例如:聚苯乙烯、聚乙烯、聚丙烯、聚酯、聚丙烯腈、苯乙烯-丁二烯共聚物、(甲基)丙烯酸酯聚合物、氟树脂、硅胶、交联葡聚糖、多糖、琼脂糖等多糖类、玻璃、金属、磁性物质、和它们的组合等。这类固相的形状可以为例如盘、球、纤维、粒子、棒、平板、容器、池、微板、试管、膜(film或membrane)、胶体、片等任意形状。具体地,可举出例如:磁性珠、玻璃珠、聚苯乙烯珠、精制琼脂糖珠、硅胶珠、多糖类珠、聚苯乙烯板、玻璃板、聚苯乙烯管等。本发明的肽固定化于这些固相可使用本领域技术人员公知的方法进行,例如,可通过物理吸附法、共价键法、离子键法等进行。固定化优选通过共价键来进行,使固相表面具有化学官能团(例如羟基、氨基、N-羟基琥珀亚胺基等),优选具有含有作为间隔体的碳原子数约4~20的亚烷基链的化学官能团,使其与肽的羧基末端进行化学反应而形成酯键或酰胺键等。固定化了本发明的肽的固相填充于亲和层析柱、HPLC柱等柱中,可以用于检测、纯化或分离人IgA。
(IgA的纯化方法)
本发明进一步提供IgA的纯化方法,该方法包括使上述本发明的肽或固定化肽与IgA结合,以及使结合的IgA游离而回收IgA。
将固定化了本发明的肽的固相填充于亲和层析柱、HPLC柱等柱中,用适当的缓冲液平衡,在室温~0℃、优选约10℃~0℃的低温(进一步优选约4℃)下施以含有人IgA的液体,使人IgA与固相上的肽结合。例如在分离血清中的IgA时,可以使用中性范围的pH、例如pH6.0~7.5的缓冲液,上柱,进行结合操作。洗脱可以在柱中流动酸性范围的pH、例如pH2~4的缓冲液(例如含有0.3M NaCl的pH3.5至pH2.5的0.2M甘氨酸-HCl缓冲液)来进行。
IgA是否已被回收可通过,例如,电泳、之后的使用抗人IgA抗体的Western印迹法来测定。电泳条件如下:可以进行使用了5~20%丙烯酰胺梯度凝胶的SDS-PAGE,另外,Western印迹条件如下:可以将电泳后的蛋白质转印至PVDF膜,用脱脂奶粉封闭后,利用抗人IgAα链山羊抗体与HRP标记抗山羊IgG小鼠抗体进行检测。
本发明的方法可用于由各种方法生成的含IgA的产物中纯化IgA的工序中获得富集IgA的级分。因此,亲和层析、HPLC等柱层析中优选使用本发明的方法。纯化IgA时,除了这类层析法之外,还可以适宜组合蛋白质的惯用纯化技术,例如,凝胶过滤层析、离子交换柱层析、反相柱层析等层析、硫酸铵分级沉淀、超滤等。
(IgA的分析方法)
本发明进一步提供IgA的检测方法,该方法包括使样品中的IgA与上述本发明的肽或固定化肽结合而检测结合的IgA。其中,检测中包含定性或定量的任一种分析。
IgA的检测可以如下进行:使用适合操作中的缓冲液,同时使样品与膜或聚苯乙烯孔板等结合,使其与本发明的标记肽接触,根据需要进行洗涤后,对标记的水平进行定性或定量。
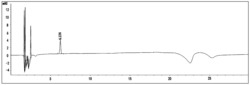
或者,在如上所述使用固定化了本发明的肽的HPLC柱时,向该柱注入含有人IgA的样品,通入结合缓冲液,使人IgA与肽结合,在例如吸光度280nm、或280nm的激发光所致的350nm的荧光下,检测蛋白质进行记录,用洗脱缓冲液(例如,含0.15M的NaCl的0.1M甘氨酸盐酸缓冲液pH2.5中的梯度洗脱)从柱洗脱,通过出现的峰和峰面积,可以进行IgA的定性和定量。
(试剂盒和柱)
本发明进一步提供用于人IgA的分析(定性、定量等)或纯化的试剂盒,其含有上述本发明的肽或固定化肽的至少1种。
本发明的试剂盒中所含的各肽或固定化肽盛放于单独的容器中。另外,根据需要,试剂盒中还可以具备记载了人IgA的分析步骤、纯化步骤的使用说明书。进而,试剂盒中还可以含有分析所需的试剂、缓冲液、固定化肽填充柱等。
本发明进而还提供IgA分离用柱,其含有上述本发明的固定化肽。
固定化肽可以将肽共价地或非共价地与通常的层析用载体(填充材料)结合来制作。这类载体的例子为:琼脂糖、精制琼脂糖等多糖类基质的载体、硅胶基质的载体、树脂基质或聚合物基质的载体等。将肽结合于载体时,可以通过烃链(例如C4~C16)之类的间隔体进行结合。
上述IgA分离用柱是用于分离IgA的柱,具体包含:用于IgA的分析或纯化?分离获得的层析柱、高效液相层析(HPLC)柱等柱。柱的尺寸没有特别限制,可以根据分析用、纯化?分离获得用等用途、施用(搭载)或注入的量等进行改变。另外,柱的材质可以可以是金属、塑料、玻璃等通常用作柱的材质。
上述的柱可以通过将根据上述手法制作的本发明的固定化肽(干燥或湿润状态)紧密地填充于柱中来制造。
实施例
以下,列举实施例进一步具体说明本发明,但本发明的范围并不受这些实施例的限制。
[实施例1]
由随机肽T7噬菌体文库分离人IgA特异性噬菌体
为了从已构建的使用了T7噬菌体展示系统的随机肽文库(CX7-10C 以及X3CX7-10CX3)分离人IgA特异性的噬菌体,使用了以下的生物淘选的方法。即,将含有0.5%BSA的PBS中的5×1010 pfu的T7噬菌体文库(CX7-10C与X3CX7-10CX3的混合物)溶液加入涂布了人IgG(多克隆,ICN/Cappel Biomedicals)(1μg/100μl/孔)并用0.5%BSA进行了封闭的96孔微板(Nunc, Maxisorp)的孔中,反应1小时(吸收步骤)。接着,将其上清转移至涂布了人IgA(来自人血浆, Athens Research &Technology, Athens, GA, USA)(1μg/100μl/孔)并用0.5%BSA(牛血清白蛋白)进行了封闭的孔中,进行1小时反应(结合步骤)。除去上清的噬菌体溶液后,用含有0.1%Tween(注册商标)的PBS(磷酸缓冲盐水)洗涤孔5次(洗涤步骤)。加入大肠杆菌5615(Novagen)的培养液(300μl)进行感染,与3ml的大肠杆菌培养液一起在37℃下孵育,直至增殖、溶菌(增殖步骤)。按照常法通过利用聚乙二醇的噬菌体沉淀法从溶菌后的培养液回收噬菌体。使所得噬菌体溶解于PBS中,通过0.45μm的过滤器后,用于随后的淘选。包含上述进行5次淘选,由此浓缩IgA特异性的噬菌体。其中,在第3~5次淘选中,在吸收步骤中使用涂布了人IgG以及小鼠IgE(各自1μg/孔)的孔,另外,在洗涤步骤中用含有0.3%Tween(注册商标)的PBS进行了10次洗涤。
相对于5次淘选后所得噬菌体的各种免疫球蛋白的结合特异性通过ELISA来调查,结果如图1所示,与使用了原来文库的ELISA的结果相比,明显可见对人IgA的结合活性的增大。
因此,将这些噬菌体随机地克隆20个克隆后,对于由ELISA观察到结合活性的10个克隆,进行展示的肽模体的分析,确定氨基酸序列(表1)。
[表1]
对于所得的肽,得到了4种模体(hA-1,2,3,4)。所得模体中,hA-2显示了最高的出现频率。另外,所得模体间的氨基酸序列的比较中显示了下述倾向:除了设计文库时导入的Cys之外,完全保留的残基为Leu5、Phe16,另外第18个优选为疏水性残基。
进行了克隆化的噬菌体(hA-1,2,3,4)对各种免疫球蛋白的结合特性的调查结果示于图2。每一克隆对人IgA均可见特异性,关于hA-2,对小鼠IgA也观察到结合活性。
为了对所得IgA特异性肽模体中的重要残基进行分析,基于结合力最高的hA-2模体,制作了如下的突变文库。即,制作了下述肽文库(文库的序列多样性为1.4×107),其中,通过混合核苷酸来合成编码图3的hIgA-2肽模体(序列编号1)的碱基序列(序列编号45)中的编码氨基酸编号4与13的Cys之外的残基的碱基,而部分地产生随机的氨基酸。混合核苷酸制作为原本的碱基序列为70%、剩余的3种碱基各为10%,结果使得原本的密码子理论上以35%的几率出现。
使用该部分突变文库,再次对hIgA进行淘选,尝试浓缩特异性的噬菌体。如图4所示,从2轮起观察到了IgA结合噬菌体的浓缩,3轮后结合活性进一步增大。因此,对3轮后的噬菌体进行克隆化,ELISA评价21个克隆的结合活性。
结果,10个克隆可见结合活性,对它们展示的模体进行了分析,结果得到了如表2所示的氨基酸序列的模体。令人感兴趣的是hA-2模体中,以氨基酸编号计,5、7、9、12号被完全保留,认为是对结合重要的残基。另外,对于氨基酸编号14与16号的残基,也被疏水性残基所占,暗示了该区域的疏水性相互作用的重要性。另一方面,对于氨基酸编号8号的残基,仅被碱性的氨基酸所占,推测存在与抗体侧的负电荷的相互作用。应予说明,上述氨基酸编号如表2所示,是16个氨基酸残基(为18氨基酸残基时,是第9个和第10个残基均缺失的剩余的16个氨基酸残基)的氨基酸编号。
[表2]
表2所示的各hA-2肽模体的氨基酸序列从上起依次(即,按照hIgA2-4, hIgA2-6, hIgA2-15, hIgA2-18, hIgA2-21, hIgA2-2, hIgA2-6, hIgA2-9, hIgA2-17, hIgA2-16的顺序)作为序列编号2, 3, 4, 5, 6, 7, 8, 9, 10, 11示于序列表中。
基于hIgA-2的肽模体来合成肽,评价该肽的结合特异性。肽使用了N末端侧添加了生物素化-(PEG)4基团的肽。在固定化了各目标蛋白质(50ng/孔)的微板的孔中,加入摩尔比为1:4的AP标记的链霉亲和素与肽的混合物(7.5nM与30nM),进行反应。用PBS-T(即,PBS-Tris)洗涤5次后,通过以PNP-磷酸为基质的发色法来检测结合。结果示于图5。关于hIgA-2肽,可知虽然存在与其它蛋白质的若干结合,但特别强地与人IgA结合。
进而,为了确认该结合的特异性,通过使用了ProteOn XPR36(Bio-Rad)的表面等离子共振分析来评价结合力(图6)。
[实施例2]
利用hIgA-2肽的人IgA的纯化
为了研究hIgA-2肽是否作为人IgA的纯化用配体发挥功能,将生物素化(PEO)4-hIgA-2固定化于1ml的HiTrap Streptavidin HP(GE Healthcare),注入各种蛋白质,评价其结合能力(图7)。作为对照蛋白质使用的人IgG、HAS(人血清白蛋白)、BSA(牛血清白蛋白)基本从柱完全通过,但IgA(5,10μg)被吸附于柱,用PBS洗涤后,通过从PBS至含有0.3M NaCl的0.2M 甘氨酸-HCl(pH2.5)的梯度洗脱而由柱洗脱。这表明hIgA-2肽具有作为人IgA纯化用配体的特异性、结合能力的基本性能。
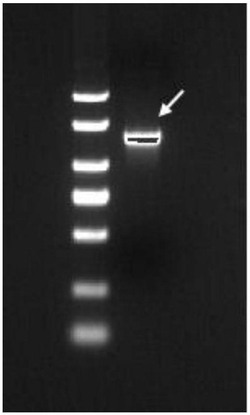
进而,为了研究hIgA-2肽固定化柱的有用性,尝试了从人血清纯化IgA。将人血清1ml用PBS稀释5倍后,上样至与Profinia蛋白质纯化系统(BioRad)连接的柱。用PBS洗涤后,通过含有0.3M NaCl的0.2M 甘氨酸-HCl (pH3.5)、接着含有0.3M NaCl的0.2M 甘氨酸-HCl(pH2.5)对结合蛋白质进行洗脱(图8)。
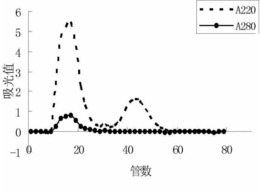
通过层析洗脱的各级分通过SDS-PAGE和使用了抗人IgA抗体的Western印迹来进行分析(图9)。酸性洗脱级分(级分D)通过SDS-PAGE后的蛋白质染色,检测到与标准品的IgA蛋白质大致相同的130-180 kDa的模糊条带。另一方面,在利用抗人IgA抗体的Western印迹中,首先,血清以及标准品的IgA,除了之前的130~180 kDa的条带之外,在300kDa以上的位置均观察到了模糊条带。认为这是,血清中的IgA虽然血清型(单体)是主要成分,但还含有一部分通过J链形成的二聚体(分泌型),因而形成上述2个条带。由柱洗脱的级分D也相同地检测到了这2个条带。由此,可知hIgA-2肽固定化柱可以用于血清型、分泌型两者的纯化。进而,作为完全通过级分的级分A或洗涤级分B、C中,基本检测不到来源于IgA的条带,因此可知该层析基本回收了血清中的IgA。
以上结果提示,包含本次分离的hIgA-2肽的类似的肽对人IgA为特异性,作为纯化用的亲和配体具有高的有用性。这种低分子(分子量约2000)的人IgA的纯化配体、检测配体尚无报道,不单作为人IgA的检测、纯化试剂有用,而且作为面向将来被期待作为新的抗体药物的人IgA抗体药物的标准纯化系统也有用。
[实施例3]
具有与hIgA显示高亲和性的A-3肽模体的肽类
由实施例2,通过使用了部分突变文库的淘选,认为由16个氨基酸组成的肽(为18个氨基酸残基时,第9个与第10个残基均缺失的剩余的16个氨基酸残基)的氨基酸编号5?7?9?12是对于与IgA结合重要的残基。因此,为了寻找显示更高亲和性的序列,与实施例1相同地制作了将上述氨基酸固定,且它们之外产生随机氨基酸的最优化文库(图10)。
使用该文库,再次进行对于hIgA的淘选,尝试了浓缩对于hIgA具有高亲和的特异性的噬菌体。如图11所示,从2轮起观察到hIgA结合噬菌体的浓缩,5轮后结合活性进一步增大。因此,克隆5轮后的噬菌体,关于结合活性优异的29个克隆的噬菌体,用ELISA评价结合特异性。
结果,得到了与A-2肽相比具有更高结合特异性和结合活性的A3-1肽(由16个氨基酸残基组成)(图12)。对显示结合活性的克隆所展示的氨基酸序列进行了分析,结果,所得模体的各氨基酸位点中,观察到了如表3所示的出现倾向。
[表3]
为了确认与hIgA的结合中的出现频率高的氨基酸残基的重要性,向A3-1肽的序列中导入了频率高的氨基酸与A-2肽中所见的氨基酸。为了同时观察各残基的侧链的重要性,进行了丙氨酸扫描。各氨基酸置换体相对于hIgA的亲和性评价通过使用Biacore T100(GE Healthcare)的表面等离子共振分析来进行。使用固定化了hIgA的CM5传感器芯片,对化学合成的各肽进行分析,基于所得的传感器曲线算出各置换体相对于hIgA的亲和性。
由部分突变文库的结果而认为重要的由16个氨基酸组成的肽(下述表4)的氨基酸编号5?7?9?12的丙氨酸(A)置换体对于IgA的结合力降低,再次确认了其重要性。其中,氨基酸编号5和7号的结合力的降低显著,认为对于与hIgA结合非常重要。另外,各置换体中,对氨基酸编号1?2?6?15?16进行了置换时,与A3-1肽相比,可见对hIgA的亲和性的提高。结果示于表4。其中,表4中的A3-1肽(A3-1, A3-1(S1R), A3-1(S1H)??A3-1(V16A), A3-1(opt1), A3-1(Opt2))的氨基酸序列各自依次作为序列编号15、16、17??42、43、44示于序列表中。
[表4]
表4中,特别是所示的肽与人IgA的结合相关的离解常数(Kd)小于1μM的肽,具有序列编号15, 16, 17, 19, 20, 24, 25, 26, 37, 38, 39, 41, 42的氨基酸序列,与人IgA具有高亲和性。
进而,基于这些结果,设计了下述肽,其在由16个氨基酸组成的肽的各氨基酸位点中导入了观察到亲和性最为提高的氨基酸。即,合成了在氨基酸编号1导入组氨酸残基?2导入蛋氨酸残基?6导入丙氨酸残基?15导入丙氨酸残基?16导入亮氨酸残基的A3-1(Opt1)肽(序列编号43)。另外,为了实现溶解性提高,合成了将氨基酸编号6和15的丙氨酸残基置换为丝氨酸残基的A3-1(Opt2)肽(序列编号44)。评价A3-1(Opt1)肽和A3-1(Opt2)肽对hIgA的亲和性,结果,各自为0.044μM和0.025μM,与A3-1相比,各自确认到约12倍和约21倍的亲和性的上升。
为了确认显示最高亲和性的A3-1(Opt2)的特异性,使用Biacore T100(GE Healthcare),向固定化了A3-1(Opt2)的传感器芯片注入hIgA1和hIgA2,由此进行表面等离子共振分析(图13)。A-2肽、A3-1肽相同地对hIgA具有高特异性,另外,评价对hIgA1和hIgA2的结合力时,可知对hIgA2的结合力稍强。
A3-1(Opt2)肽是与A-2肽相比具有约52倍亲和性的肽。因此,制作A3-1(Opt2)肽柱,进行从人血清纯化hIgA。将人血清1ml用PBS稀释5倍后,上样至与Profinia蛋白质纯化系统(BioRad)连接的柱。用PBS洗涤后,用0.2M 甘氨酸-HCl (pH2.5)洗脱结合蛋白质(图14)。与A-2肽柱相比(图8),洗脱级分中检测到更为明确的峰。通过亲和性的提高,推测吸附于柱内的hIgA量有所增加。
产业实用性
本发明提供可以与人IgA特异性或选择性地结合的肽,由此,在产业上可以用于作为抗体药物的IgA的制造中的IgA的纯化,以及IgA的分析。
本说明书中引用的全部发行刊物、专利和专利申请作为参考全部引入本说明书中。
序列表正文
序列编号1~44:IgA结合性肽
序列编号45:编码IgA结合性肽A-2的DNA
序列表
<110> Kagoshima University
Otsuka Chemical Co., Ltd.
<120> IgA结合性肽以及利用其的IgA的纯化
<130> PH-4823-PCT
<150> JP 2010-118508
<151> 2010-05-24
<160> 45
<170> PatentIn 3.4版
<210> 1
<211> 16
<212> PRT
<213> 人工
<220>
<223> IgA结合性肽
<400> 1
His Met Arg Cys Leu His Tyr Lys Gly Arg Arg Val Cys Phe Leu Leu
1 5 10 15
<210> 2
<211> 16
<212> PRT
<213> 人工
<220>
<223> IgA结合性肽
<400> 2
Gln Met Arg Cys Leu Ser Tyr Lys Gly Arg Arg Val Cys Leu Trp Leu
1 5 10 15
<210> 3
Q:办理专利转让的流程及所需资料
A:专利权人变更需要办理著录项目变更手续,有代理机构的,变更手续应当由代理机构办理。
1:专利变更应当使用专利局统一制作的“著录项目变更申报书”提出。
2:按规定缴纳著录项目变更手续费。
3:同时提交相关证明文件原件。
4:专利权转移的,变更后的专利权人委托新专利代理机构的,应当提交变更后的全体专利申请人签字或者盖章的委托书。
Q:专利著录项目变更费用如何缴交
A:(1)直接到国家知识产权局受理大厅收费窗口缴纳,(2)通过代办处缴纳,(3)通过邮局或者银行汇款,更多缴纳方式
Q:专利转让变更,多久能出结果
A:著录项目变更请求书递交后,一般1-2个月左右就会收到通知,国家知识产权局会下达《转让手续合格通知书》。









动态评分
0.0