

IPC分类号 : C12N9/10,C12N15/54,C12N15/63,C12N5/10,C12N1/21,C12P5/00,C12P7/04,A01N31/02,A01N27/00,A01P17/00,C12N15/82,A01H5/00,C12R1/19








专利摘要
本发明公开一种与倍半萜合成相关的蛋白及其编码基因与应用。本发明提供的蛋白,是如下1)或2)的蛋白质:1)由序列表中序列2所示的氨基酸序列组成的蛋白质;2)将序列表中序列2的氨基酸序列经过一个或几个氨基酸残基的取代和/或缺失和/或添加且与倍半萜合成和/或驱避蚜虫相关蛋白的由1)衍生的蛋白质。本发明的实验证明,本发明得到普通小麦Tafps-B1cDNA的核酸序列及Tafps-B1编码蛋白序列;大肠杆菌中表达的TaFPS-B1蛋白具有FPP合成活性;采用农杆菌介导法体获得转小麦Tafps-B1基因拟南芥,结果表明转Tafps-B1的植株可显著驱避蚜虫。
说明书
技术领域
本发明涉及生物技术领域,尤其涉及一种与倍半萜合成相关的蛋白及其编码基因与应用。
背景技术
桃蚜是广食性害虫,寄主约有350多种,主要危害烟草、桃、李、梅、梨等果树和白菜、萝卜、辣椒、菠菜等蔬菜,造成了很大的经济损失。近年来,由于全球气候变暖、耕作制度变化等因素,使蚜虫的繁殖能力和适应性显著增强,其危害日趋严重。目前,蚜虫防治以喷洒农药为主,但大量使用农药,不仅对人畜有害,而且造成了严重的环境污染。培育抗蚜虫小麦和蔬菜品种是防止蚜虫危害的最有效途径,但由于现有种质资源中缺乏有效的抗蚜基因,抗性机制尚不明确,常规育种难以奏效。挖掘和利用新型抗蚜基因并通过基因工程培育小麦、棉花和蔬菜抗蚜新种质具有重要意义。
倍半萜类化合物在植物与植食性昆虫互作中发挥重要作用,参与植物的防御反应。法尼烯焦磷酸合成酶(farnesyl pyrophosphate synthase,FPS)是萜类化合物合成途径中的关键酶之一,其催化合成的产物FPP可在细胞质中通过MVA(Mevalonate pathway)途径作为[反]-β-法尼烯【(E)-β-farnesene,EβF】合成酶的底物。EβF是重要的倍半萜类化合物,是一种无色无味挥发物,是小麦麦长管蚜(Sitobion avenae Fabricius)、禾谷缢管蚜(Rhopalosi phum padi Linnaeus)、麦无网长管蚜(Metopolophium dirhodum Walker)和麦二叉蚜(Schizaphis graminum Rondani)等蚜虫报警信息素的唯一成份,可以使蚜虫产生骚动、从植株上脱落,并吸引蚜虫天敌,有效控制蚜虫危害。研究发现,许多植物中含有EβF合成酶,能释放EβF,具有天然的蚜虫驱避特性。但小麦、棉花、大豆等重要农作物中,只含有FPS,但不含有EβF合成酶。Yu等发现转EβF合成酶烟草表明EβF的释放量受前体物质FPP供应量的限制。
目前已有20多种植物的FPS编码基因得到分离,但其在其它植物的表达和功能研究只有以下两例报道。Chen等利用农杆菌介导法所获得的转棉花fps基因的黄花蒿(Artemisia annua)的倍半萜生物合成途径的代谢流明显增加,使抗疟类药物—青蒿素增加明显,部分株系比野生株系高4倍。崔红等将薄荷fps基因导入烟草发状根,抑菌试验发现转基因发状根汁液对赤星病菌具有抗性。小麦fps基因的分离克隆、表达特性、酶活特性和在植物中(拟南芥)的表达与抗蚜虫功能研究尚无报道。
发明内容
本发明的一个目的是提供一种与倍半萜合成相关的蛋白及其编码基因。
本发明提供的蛋白,是如下1)或2)的蛋白质:
1)由序列表中序列2所示的氨基酸序列组成的蛋白质;
2)将序列表中序列2的氨基酸序列经过一个或几个氨基酸残基的取代和/或缺失和/或添加且与倍半萜合成和/或驱避蚜虫相关蛋白的由1)衍生的蛋白质。
上述(a)或(b)中的蛋白可人工合成,也可先合成其编码基因,再进行生物表达得到。上述(b)中的蛋白的编码基因可通过将序列表中序列1所示的DNA序列缺失一个或几个氨基酸残基的密码子,和/或进行一个或几个碱基对的错义突变,和/或在其5′端和/或3′端连上表2所示的标签的编码序列得到。
上述蛋白的氨基酸序列中一个或几个氨基酸残基的取代、替换和/或添加,有的是由于自然发生的变异引起的,有的是由人工诱变处理引起的。
上述倍半萜为法尼醇和/或β-石竹烯;上述蚜虫为桃蚜。
上述蛋白的编码基因也是本发明保护的范围。
上述编码基因为为1)-4)中任一一种:
1)序列表中序列1所示的DNA分子;
2)序列表中序列1自5’末端第650-1129位核苷酸所示的DNA分子;
3)在严格条件下与1)或2)限定的DNA序列杂交且编码与倍半萜合成和/或驱避蚜虫相关蛋白的DNA分子;
4)与1)或2)限定的DNA序列至少具有70%、至少具有75%、至少具有80%、至少具有85%、至少具有90%、至少具有95%、至少具有96%、至少具有97%、至少具有98%或至少具有99%同源性且编码与倍半萜合成和/或驱避蚜虫相关蛋白的DNA分子。
上述严格条件为在6×SSC,0.5%SDS的溶液中,在65°C下杂交,然后用2×SSC,0.1%SDS和1×SSC,0.1%SDS各洗膜一次。
扩增上述基因全长或其任意片段的引物对也是本发明保护的范围。
含有上述编码基因的重组载体或转基因细胞系或重组菌也是本发明保护的范围。
上述重组载体为将上述编码基因插入到表达载体中,得到表达所述蛋白的载体。
上述重组载体具体为如下1)或2):
1)将序列表中序列1插入载体pEASY-E1的EcoRⅤ和BstXⅠ酶切位点间得到的载体;
2)将序列表中序列1插入载体pBI121的Xma I和Sac I间得到的载体。
上述重组菌为将上述的1)所示的重组载体导入目的菌得到的重组菌;所述目的菌具体为大肠杆菌。
上述蛋白或上述编码基因或上述重组载体或上述的重组菌在合成倍半萜和/或驱避蚜虫中的应用也是本发明保护的范围。
上述应用中,上述倍半萜为β-石竹烯和/或法尼醇;上述蚜虫为桃蚜。
本发明的第二个目的是提供一种培育释放倍半萜转基因植物的方法。
本发明提供的方法,为将上述蛋白的编码基因导入目的植物得到转基因植物,所述转基因植物的倍半萜释放量高于所述目的植物;
上述蛋白的编码基因具体通过上述2)所示的重组载体导入目的植物;
上述倍半萜具体为β-石竹烯;
上述目的植物具体为双子叶植物或单子叶植物,上述双子叶植物进一步具体为拟南芥。
本发明的第三目的是提供一种培育抗蚜虫转基因植物的方法。
本发明提供的方法,为将上述蛋白的编码基因导入目的植物得到转基因植物,所述转基因植物的抗蚜虫能力高于所述目的植物;
上述倍半萜具体为β-石竹烯;
上述蛋白的编码基因具体通过上述2)所示的重组载体导入目的植物;
上述目的植物具体为双子叶植物或单子叶植物,上述双子叶植物进一步具体为拟南芥。
本发明的实验证明,本发明得到普通小麦Tafps1基因型中的Tafps-B1cDNA的核酸序列及Tafps-B1编码蛋白序列,桃蚜可诱导苗期小麦Tafps-B1基因在根中表达量降低,在茎表达量升高,其中在叶片中表达量显著升高(P<0.05);抽穗期小麦Tafps-B1基因在叶片(P<0.01)和茎(P<0.05)中的表达量显著升高,在幼胚中轻微升高,在颖壳中显著降低(P<0.05),根中轻微降低,推测该基因与小麦对蚜虫侵害的应激抗性反应相关,具有防御反应诱导的表达特性。利用大肠杆菌中表达的TaFPS-B1蛋白;发现其可以具有FPP合成活性,可催化前体化合物异戊基焦磷酸(isopentenyl pyrophosphate,IPP)和香叶草基焦磷酸(geranyl pyrophosphate,GPP)合成法尼基焦磷酸(farnesyl pyrophosphate,FPP);采用农杆菌介导法体获得转Tafps-B1拟南芥,GC及GC-MS分析表明苗期转Tafps-B1的拟南芥倍半萜β-石竹烯的释放量是野生型的5.5倍;4通道嗅觉仪检测有翅桃蚜对花期转Tafps-B1的拟南芥挥发物的反应表明转Tafps-B1的植株对蚜虫有极其显著得驱避作用(P<0.01),与野生型拟南芥相比,转Tafps-B1的植株可显著驱避蚜虫。表明普通小麦Tafps-B1的编码区序列可应用于通过转基因技术提高小麦、棉花和蔬菜等农作物抗蚜性的研究。
附图说明
图1为普通小麦Tafps-B1cDNA的PCR扩增
图2为普通小麦接种蚜虫后Tafps1基因表达量的变化
图3为Tafps-B1基因体外酶化反应产物法尼醇的鉴定
图4为转Tafps-B1的拟南芥的分子检测
图5为GC及GC-MS对苗期转Tafps-B1的拟南芥β-石竹烯的释放量的比较分析
图6为4通道嗅觉仪检测有翅桃蚜对花期转Tafps-B1的拟南芥挥发物的取食反应的比较图
具体实施方式
下述实施例中所使用的实验方法如无特殊说明,均为常规方法。
下述实施例中所用的材料、试剂等,如无特殊说明,均可从商业途径得到。
下述实施例中野生型拟南芥Col-0型(Cunillera N,Arro M,Delourme D,Karst F.,Boronat A.,FerrerA.1996,Arabidopsis thaliana contains two differentially expressed farnesyl-diphosphate synthase genes.Journal of Biological Chemestry271(13):7774-7780.公众可从中国农业科学院作物科学研究所获得)种植于中国农业科学院作物科学研究所人工气候箱中(温度(22±2)℃,湿度60%—80%,光周期L∶D=16∶8)。
下述实施例中桃蚜记载Beale MH,Birkett MA,Bruce TJA,Chamberlain K,Field LM,Huttly AK,Martin JL,Parker R,Phillips AL,Pickett JA,Prosser IM,Shewry PR,Smart LE,Wadhams LJ,Woodcock CM,Zhang YH.2006.Aphid alarm pheromone produced by transgenic plants affects aphid and parasitoid behavior.Proc Natl Acad Sci USA,103(27),10509-10513,公众可从中国农业科学院作物科学研究所获得;将蚜虫接种到拟南芥植株上(人工饲养),温度(20±2)℃,湿度60%80%,光周期L∶D=16∶8。
下述实施例中质粒提取试剂盒购自Biomega公司,内切酶Xma I和Sac I购自NEB公司,高保真酶pfu、T-easy载体、大肠杆菌菌株DH5α、反转录试剂盒购自北京全式金公司,rTaq DNA聚合酶购自TaKaRa公司,总RNA的提取试剂盒购自天根生化科技有限公司,去磷酸酶Alkaline Phosphatase购自Promega(Catalog:M1821),底物IPP(Product Number:I0503)和GPP(Product Number:G6772)以及正己烷等其他化学试剂均购自sigma公司,其他化学试剂为国产分析纯,引物和测序均由北京华大基因完成。
实施例1、普通小麦Tafps-B1cDNA的获得
一、普通小麦Tafps-B1cDNA的获得
1、普通小麦总RNA的提取和cDNA的合成
普通小麦的总RNA的提取参照天根生化科技有限公司RNA simple总RNA提取试剂盒中的说明手册提取。琼脂糖凝胶电泳检测并通过NanoDrop测定浓度。
cDNA第一链的合成采用北京全式金生物技术有限公司的TransScript First-Strand cDNA Synthesis SuperMix试剂盒。方法参照试剂盒中的说明手册。具体操作如下:
第一链cDNA合成
1)加入
2)轻轻混匀,42°C孵育30min。
3)85°C加热5min失活TransScript RT Enzyme。反应合成的cDNA第一链作为PCR反应的模板待用,得到cDNA。
2、引物设计与基因克隆
根据拟南芥Atfps1基因(At5G47770.1),在TIGR注释的水稻基因组网站(http://rice.plantbiology.msu.edu/)上,选择相似性大于80%的,查找结果有Os05g46580和Os01g50760。以水稻Os01g50760(Ofps1)序列在TIGR注释的植物转录装配数据库中(http://plantta.jcvi.org/index.shtml)选择wheat进行blast,获得小麦fps1基因序列。DNAman5.0比对显示保守区域存在于5′UTR和3′UTR。分别在5′UTR及3′UTR处设计保守引物Full-fps1F:5′-TACTCTCCACAGAAATCTCCGCTC-3′,Full-fps1R:5′-CAACAACCTCTCCAAGAGCTTAGC-3′,由北京华大基因公司合成。
PCR反应体系为5×Tans FastPfu Buffer10μl、dNTP(2.5mmol·l-1)5μl、Tans FastPfu DNA Polymerase 1μl,Forward primer(10μmol·l-1)1μl、Reverse primer(10μmol·l-1)1μl、cDNA1μl,用ddH2O补足至50μl。PCR的反应条件为95℃2min;95℃20s,60℃20s,72℃15s,40个循环;72℃5min;4℃保存。
普通小麦科农199记载李俊明,张相岐,张爱民,王志国,安调过,纪军,王静.高产广适小麦新品种科农199,2007.麦类作物学报,27(2),368.普通小麦科农199,公众可从中国农业科学院作物科学研究所获得。
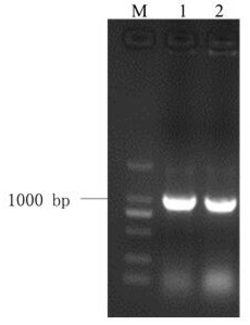
以普通小麦科农199的cDNA为模板,用Full-fps1F/R作为引物进行扩增,得到1154bp PCR产物(含64bp的5′UTR和25bp的3′UTR序列),将PCR产物电泳,结果如图1所示,M:Trans2K DNA marker;1、2:PCR产物,可以看出得到1154bp目的片段。目的片段利用TA克隆的方法连接到T-easy载体(购自北京全式金公司)后,构建成T-Tafps-B1测序载体,经过测序,该1154bp PCR产物包含1065bp的基因编码区,该1154bp PCR产物的核苷酸序列为序列表中的序列1,其中含有基因的编码区位于第65-1129位,将该基因命名为Tafps-B1,该基因编码的蛋白命名为TaFPS-B1;该蛋白的氨基酸序列为序列表中的序列2,共354个氨基酸。经过序列比对,Tafps-B1与拟南芥Atfps1核苷酸的相似性为61.13%。Tafps-B1与水稻Osfps1核苷酸的相似性为87.98%。
T-Tafps-B1测序载体为将序列表中的序列1插入T-easy载体得到的载体。
二、普通小麦接种蚜虫后Tafps-B1基因表达量的变化
普通小麦科农199苗期材料选择种子发芽后培养3周二叶期的材料;抽穗期选择小麦开花后12d,幼胚刚形成的时期的材料。
苗期接种桃蚜虫30头,4d后取对照与接种桃蚜虫植株的根、茎和叶的器官,提取RNA,反转录得到cDNA;
抽穗期接种桃蚜虫100头,4d后取对照与接种蚜虫植株的根、茎、叶、幼胚的器官,提取RNA,反转录得到cDNA。
分别利用两对引物taqrtfps1.1F:5′-TCCTTCAACGCCACTTCAGAGGAA-3′,taqrt-fps1.1R:5′-TGTCTTGTATTGCACAATGCGCCG-3′和taqrt-fps1.2F:5′-ATCCTTCAACGCCACTTCAGAGGA-3′,taqrt-fps1.2R:5′-TGTCTTGTATTGCACAATGCGCCG-3′进行real-time quantitative PCR的检测分析。
具体实施方法见上述一中的普通小麦总RNA的提取和cDNA的合成,在提取幼胚及种子RNA时将天根试剂盒中的RZ溶液换为HZ溶液。
结果见图2所示,白色柱:接种前;黑色柱:接种后;**表示经t测验接种蚜虫后的表达量与接种前的差异极显著(P<0.01);*表示经t测验接种蚜虫后的表达量与接种前差异显著(P<0.05),其中,图2A为Tafps-B1在苗期接种蚜虫前后不同器官表达量变化;图2B为Tafps-B1在抽穗期接种蚜虫前后不同器官表达量变化;可以看出,接种蚜虫后,苗期小麦Tafps-B1基因在根中表达量降低约46%,在茎中上升约52%,在叶片中显著上升,约100%(P<0.05),见图2A;抽穗期小麦Tafps-B1基因在叶片(P<0.01)和茎(P<0.05)中的表达量显著升高,分别为209%和63%。在幼胚中升高125%(P=0.15),在颖壳中显著降低(P<0.05),为62%,根中轻微降低,67%(P=0.32),见图2B。推测该基因与小麦对蚜虫侵害的应激抗性反应相关,具有防御反应诱导的表达特性。
实施例2、Tafps-B1的功能鉴定
一、Tafps-B1在合成倍半萜化合物法尼醇中的应用
1、载体构建
以T-Tafps-B1测序载体为模板,利用高保真酶TranStart FastPfu DNA Polymerase及引物efpps1F(5′-ATGGCGGCGGCGGCGGTGGCGTTG-3′)/efps1R(5′- CTATTTCTGCCTCTT-3′,加粗斜体引物部分是3′UTR序列),得到1074bp的带有A末端的目的片段(具有序列表中序列1的核苷酸)。将上述带有A末端的目的片段参照全式金生物有限公司pEASY-E1 Expression Kit说明书与载体pEASY-E1连接,得到的连接产物转入普通大肠杆菌后涂于含Amp(50mg/ml)的LB平板上过夜培养。
挑取单克隆利用T7F(5′-TAATACGACTCACTATA-3′)及目的基因下游引物efps1R进行检测,得到1216bp的阳性克隆。将阳性克隆提取质粒送去测序,该质粒为将序列表中序列1自5’末端第65-1129位核苷酸利用TA克隆的方法插入载体pEASY-E1的EcoRⅤ和BstXⅠ酶切位点酶切位点间得到的载体(6个组氨酸后24bp),命名为pEASY-E1-Tafps-B1。
将pEASY-E1-Tafps-B1转化BL21(DE3)感受态细胞,转化过程见全式金生物有限公司BL21(DE3)Chemically Competent Cell说明书,得到BL21(DE3)/pEASY-E1-Tafps-B1,将BL21(DE3)/pEASY-E1-Tafps-B1利用T7F及目的基因下游引物进行PCR检测,得到1216bp的目的片段;证明pEASY-E1-Tafps-B1已转入大肠杆菌中。
采用同样的方法将空载体pEASY-E1转入BL21(DE3)中,得到BL21(DE3)/pEASY-E1。
2、TaFPS-B1蛋白的诱导表达、分离及纯化
将BL21(DE3)/pEASY-E1-Tafps-B1在加入1μM的IPTG的LB培养基中培养,条件为16℃180rpm、诱导12h,得到培养产物,将培养产物于4℃、1000rpm离心10min收集上清液。
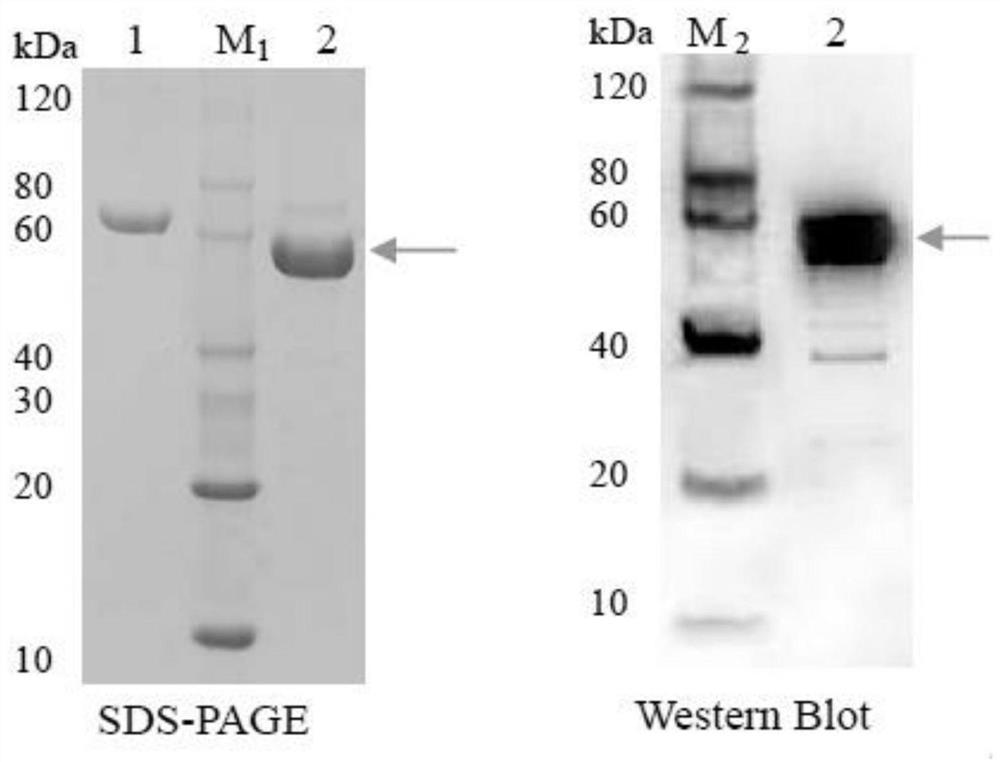
将上清液利用 FPLC system(Amersham),HiLoad_16/60_superdex_75_prep_grade(global)分子筛预装柱分离纯化(4ml上样环,洗脱流速:1ml/min,每1ml收集一次;洗脱液(50mM Na2HPO4,300mM NaCl,250mM imidazole,NaOH调pH至8.0),经SDS-PAGE检验后获得46KD纯化TaFPS-B1蛋白(为收集2柱体积的洗脱液)。
将BL21(DE3)/pEASY-E1采用上述方法诱导、分离及纯化,未得到46KD目的蛋白。
3、TaFPS-B1蛋白酶活性鉴定
利用蛋白超滤膜离心管,将收集2柱体积的洗脱液(纯化TaFPS-B1蛋白)用pH为7.0、50mM PBS(通过将39ml0.2M的NaH2PO4与61ml0.2M的Na2HPO4混匀后获得)置换,并将蛋白浓缩4μg/ml。为防止有机溶剂的污染,以下均采用玻璃器皿。按照表1的体系进行反应,体系1mL,37°C,30min,得到反应产物。以水替代纯化TaFPS-B1蛋白作为对照。
表1为TaFPS蛋白酶活性鉴定反应体系
鉴于产物应该为法尼烯焦磷酸(FPP),难以检测及定量,因此利用去磷酸酶Alkaline Phosphatase购自Promega(Catalog:M1821)对产物进行去磷酸:
具体反应体系为CIAP10×Reaction Buffer6,CIAP(1U/)6μl,ddH2O48μl,混匀后分别取20μl加入各反应后的样品中,37°C孵育30min;重复以上步骤。
反应结束后,每个样品瓶子中加入500μl的正己烷,涡旋1min,冰上放置30min,抽提产物。小心吸取上清至新的玻璃瓶中,利用稳定的氮气流(14PSI)吹到只剩2μl(约5min),加入10μl的正己烷,待用。
取1μl样品进样,利用气质联用系统GC-MS(HP6890/HP5973,Hewlett-Packard公司),色谱柱HP-5MS(60m×250μm×0.25μm;Hewlett-Packard公司),GC-MS仪器进行分析。升温程序为:40℃保持1min,以5min5℃的增量升到250℃(1℃/min),最后250℃保持17min。MS检测范围为40–550Da,温度为170℃。
GC-MS检测两组的反应产物,结果如图3所示,(A)对照组样品的GC-MS检测结果;(B)对照组中35.003min峰的成分鉴定,鉴定结果为溶剂而非法尼醇;(C)纯化TaFPS-B1蛋白GC-MS检测结果,主峰为法尼醇;(D)纯化TaFPS-B1蛋白中34.911min特异峰的质谱图;(E)数据库中法尼醇的质谱图及化学结构式;可以看出,在对照组中未检测到法尼醇(图3A;图3B);在纯化TaFPS-B1蛋白样品的34.911min时检测到主峰(图3C)将其进行质谱分析,该主峰为法尼醇(图3D),与数据库中的法尼醇质谱值相匹配(图3E)。
说明纯化TaFPS-B1蛋白可催化前体化合物异戊基焦磷酸(isopentenyl pyrophosphate,IPP)和香叶草基焦磷酸(geranyl pyrophosphate,GPP)合成法尼基焦磷酸(farnesyl pyrophosphate,FPP),并通过去磷酸化形成法尼醇。
二、Tafps-B1在合成倍半萜化合物β-石竹烯中的应用
1、转Tafps-B1拟南芥的获得
1)、表达载体的构建
设计带有Xma I和Sac I酶切位点的目的基因引物XSfps1F:5′-TCCCcccgggATGGCGGCGGCGGCGGTG-3′(小写及下划线标注Xma I位点),XSfps1R:5′-ACgagctcAGCTTAGCTCTATTTCTGCCTCTT-3′(小写及下划线标注Sac I位点,AGCT为3′UTR序列,为协调引物的GC含量和退火温度),以普通小麦科农199的cDNA为模板,利用高保真酶进行扩增。PCR反应体系为5×Tans FastPfu Buffer 10μl、dNTP(2.5mmol·1-l-1)5μl、Tans FastPfu DNAPolymerase 1μl,Forward primer (10μmol·L-1)1μl、Reverse primer(10μmol·l-1)1μl、cDNA1μl,用ddH2O补足至50μl。PCR的反应条件为95℃2min;95℃20s,60℃20s,72℃15s,40个循环;72℃5min;4℃保存。
1%琼脂糖凝胶电泳,结果为得到1087bp的PCR扩增产物,切胶回收,得到回收片段。将pBI121(王华新,曹家树,向珣,余小林,叶纨芝.2008,pBI121表达载体及其转化植株的快速鉴定.浙江大学学报:农业与生命科学版,28(2),137-142.公众可从中国农业科学院作物科学研究所获得)过表达载体质粒与回收片段同时利用Xma I和Sac I进行4h双酶切,酶切体系为10×NEB Buffer45μl,100×BSA0.5μl,pBI121质粒或回收片段2μg,Xma I1μl,Sac I1μl,用ddH2O补足至50μl。纯化回收酶切产物。T4连接酶4℃过夜连接,连接体系为pBI121片段3μl,目的基因酶切片段2μl,10×Buffer 1μl,T4DNA Ligase0.5μl,用ddH2O补足至10μl。连接产物转化大肠杆菌后,得到转化子。提取转化子的质粒,用引物XSfps1F/R和XSfps2F/R进行PCR检测、酶切鉴定,将PCR和酶切均得到1087bp的质粒送测序,测序结果表明该质粒为将序列表中序列1自5’末端第65-1129位核苷酸插入pBI121的Xma I和Sac I酶切位点得到的质粒,将该质粒命名为pBI121-Tafps-B1。
2)、农杆菌介导的拟南芥转化(浸花法)
A:农杆菌电击感受态的制备
(1)挑取农杆菌AGL1(He Y,Jones HD,Chen S,Chen XM,Wang DW,Li KX,Wang DS,Xia LQ.2010,Agrobacterium-mediated transformation of durum wheat(Triticum turgidum L.var.durum cv Stewart)with improved efficiency.Journal of Experimental Botany,61(6):1567–1581.公众可从中国农业科学院作物科学研究所获得)单菌落接种到1ml LB液体培养基中,短暂涡旋使细胞充分悬浮,将悬浮液接种到10ml含羧苄霉素400mg/L的LB液体培养基中,28℃,250rpm震荡培养过夜;
(2)将10ml已培养的菌液接种到100ml含400mg/L羧苄霉素的LB液体培养基中,28℃,250rpm震荡培养,在OD值为0.5左右时,进行操作;
(3)将在冰上静置30min后菌液的,在4℃条件下,4000rpm离心10min,去上清,加入20ml10%的甘油重悬菌体;
(4)在4℃条件下,4000rpm离心10min,弃上清,加入10ml含有10%甘油重悬菌体;
(5)在4℃条件下,4000rpm离心10min,弃上清,缓慢加入2ml含有10%甘油重悬菌体;
(6)将制好的感受态分装(每管50μl),液氮冷冻后于-80℃保存备用。
B:电击转化农杆菌
(1)在冰上将1μl的重组质粒pBI121-Tafps-B1加入50μl电击农杆菌感受态中,轻轻混和均匀;
(2)将混合物加入电极杯的中央后再把电极杯放入槽孔中;
(3)调农杆菌电击参数,电击混合物;
(4)加入100μl LB液体培养基后将混合物吸出,再加入500μl LB液体培养基,28℃,230rpm复苏2h;
(5)将上述混合物均匀涂在含100mg/L Kan和100mg/L Carb的LB平板上;
(6)在28℃黑暗温箱中倒置培养2~3d;
(7)挑取单克隆菌株进行PCR检测。
C:浸花法拟南芥转化
1ml:20ml接种农杆菌。28℃,250rpm过夜培养,次日按5ml:100ml接种,28℃,250rpm过夜培养,次日上午测OD值,用LB+Kan+Carb作为空白对照,当菌液达到OD600为0.8左右时,4℃,5000g离心12min收集菌体。100ml悬浮液(1/2MS,10mM MgCl2,5%蔗糖,500μl/L Silwelt)悬浮后,导入小烧杯中用于转化。转化时将野生型拟南芥的花在溶液中浸泡1min左右,浸泡结束后,将pBI121-Tafps-B1放倒并将每个花枝与其他花枝分开,避光保温保湿过夜。隔周再次侵染,一共侵染4次,得到T0代转Tafps-B1拟南芥种子。
3)、转Tafps-B1拟南芥的筛选及分子鉴定
将2000个T0代转Tafps-B1拟南芥种子消毒灭菌后,点种于MS+Kan(50mg/L)平板筛选,获得3棵T0代转Tafps-B1拟南芥。
从T0代转Tafps-B1拟南芥收获种子、播种,得到T1代转Tafps-B1拟南芥。
提取T1代转Tafps-B1拟南芥的cDNA为模板,利用Jcfps1F:5′-TTTAGGGTGCCTCAGGTTGGCCTT-3′,Jcfps1R:5′-GACAGCAGCAATGCACATGCAACC-3′为引物对,进行PCR扩增;
电泳结果如图4A所示,M:Trans2K DNA marker;WT为野生型拟南芥,1-3为T1代转Tafps-B1拟南芥,得到296bp为阳性T1代转Tafps-B1拟南芥,表明Tafps-B1已转到野生型拟南芥中。T1代植株进行PCR鉴定表明凡是经过筛选成活的全被检测为阳性。
从T1代转Tafps-B1拟南芥收获种子、播种,得到T2代转Tafps-B1拟南芥。
提取T2代转Tafps-B1拟南芥不同组织器官的cDNA为模板,利用引物taqrt-fps1.1F:5′-TCCTTCAACGCCACTTCAGAGGAA-3′,taqrt-fps1.1R:5′-TGTCTTGTATTGCACAATGCGCCG-3′为引物对,进行qRT-PCR扩增;
以拟南芥18S rRNA基因为内参,所用引物为At18sRNA F:5′-GAAACGGCTACCACATCCA-3′,At18sRNAR:5′-ATCCCAAGGTTCAACTACGAG-3′。
在ABI PRISM7900实时荧光定量PCR仪升进行操作,设3次重复。
结果如图4B所示,T-WT-R:T2代转Tafps-B1拟南芥根;T-WT-S:T2代转Tafps-B1拟南芥茎;T-WT-L:T2代转Tafps-B1拟南芥叶;T-WT-F:T2代转Tafps-B1拟南芥花;T-WT-SL:T2代转Tafps-B1拟南芥荚;可以看出,Tafps-B1在转基因拟南芥的根(R,Root)、茎(S,Stem)、叶(L,Leaf)、花(F,Flower)和夹(SL,siliques)中都有表达。
采用同样的方法将空载体pBI121转入野生型拟南芥中,得到T0代转空载体拟南芥,播种传代直到得到T2代转空载体拟南芥,用T1代转Tafps-B1拟南芥的鉴定方法,结果没有目的片段,说明T2代转空载体拟南芥为阳性T2代转空载体拟南芥。
2、转Tafps-B1拟南芥的产生倍半萜β-石竹烯
A:拟南芥的气体收集
为了避免土壤中挥发物的影响,利用ddH2O将土洗净后,将植物放入玻璃瓶中,并灌满ddH2O以保证植物正常生长。利用顶端开小口的玻璃容器、密闭的铝质金属底座以及金属夹子组装成相对封闭的系统,对拟南芥进行气体收集。按照密闭的原则组装装置,使气体利用真空泵通过炭柱过滤后进入装置,含有吸附剂的玻璃管连接于出气口。进气的流速(600ml/min)略大于出气的流速(400ml/min)。
选择苗期(播种后第28天,4周)T2代转Tafps-B1拟南芥进行气体采集。鉴于拟南芥植株较小,挥发物较少,吸附剂在开花前期采用50mg Tenax TA聚合物,开花后采用50mg PorapaK Q聚合物。Tenax玻璃管使用前利用5ml重蒸乙醚清洗3次后,在持续的氮气通过的条件下220℃加热2h;PorapaK Q玻璃管则使用前利用1mL超纯乙醚清洗3次以上后,在持续的氮气通过的条件下132℃加热2h。气体收集完毕后,Tenax玻璃管直接用于GC-MS分析,PorapaK Q玻璃管则需要利用250μl重蒸乙醚洗脱2次(共500μl)至安捷伦玻璃瓶中,待用。以苗期野生型拟南芥、苗期T2代转空载体拟南芥为对照。
分别得到野生型拟南芥挥发物样品、T2代转空载体拟南芥挥发物样品和T2代转Tafps-B1拟南芥挥发物样品。
B:GC及GC-MS对苗期转Tafps-B1的拟南芥β-石竹烯的释放量的比较分析
用GC(Agilent公司6890N,色谱柱参数:长50m×内径0.32mm×薄膜厚度0.52μm的非极性色谱柱;电离参数:70eV,220℃;气相色谱炉温度保持在30℃1分钟,然后编程在5℃/min升温至150℃,接着10℃/min升温至150℃。)对野生型拟南芥挥发物样品、T2代转空载体拟南芥挥发物样品和T2代转Tafps-B1拟南芥挥发物样品进行检测,
结果如图5A和5B所示,其中,5A为野生型拟南芥挥发物样品GC峰图,5B为T2代转Tafps-B1拟南芥挥发物的GC峰图;可以看出,T2代转Tafps-B1拟南芥挥发物明显增多(图5B)。
用GC-MS(Agilent公司6890N,色谱柱参数:长50m×内径0.32mm×薄膜厚度0.52μm的非极性色谱柱;电离参数:70eV,220℃;气相色谱炉温度保持在30℃5分钟,然后编程在每分钟5℃升温至250℃摄氏度。)对上述GC峰图中的各组分主要的峰进行鉴定与分析,结果为主要有壬醛(Nonanal,图5A和图5B的1)、癸醛(Decanal,图5A和图5B的3)、异十三碳醛(Tridecanal,图5A和图5B的4)、β-石竹烯(Caryophyllene,图5A和图5B的5)、戊酸(Pentanoic acid,图5A和图5B的6)。
可以看出,β-石竹烯为倍半萜,因此下面对此化合物进行详细研究鉴定:
收集T2代转Tafps-B1拟南芥挥发物样品停留时间为35.889min的GC峰(图5B的5)进行质谱鉴定,方法同上,结果如图5C所示,说明该停留时间为35.889min的GC峰对应的样品为β-石竹烯,说明T2代转Tafps-B1拟南芥挥发物含有倍半萜β-石竹烯。
将人工合成的标准样品β-石竹烯(可从上海香杰香精香料有限公司购买到)进行GC(Agilent公司6890N,色谱柱参数:长50m×内径0.32mm×薄膜厚度0.52μm的非极性色谱柱;电离参数:70eV,220℃;气相色谱炉温度保持在30℃1分钟,然后编程在5℃/min升温至150℃,接着10℃/min升温至150℃)检测,结果如图5D所示,主峰的保留时间为27.815min。由于GC和GC-MS仪器中所使用的程序和参数不同,利用人工合成的100ng/μlβ-石竹烯标准样品在GC仪器中的停留时间为27.318min,且峰面积也不同。
数据库中β-石竹烯的质谱图及化学结构式如图5E所示,其中质谱图与图5C一致,进一步证明,收集T2代转Tafps-B1拟南芥挥发物样品停留时间为35.889min的样品为β-石竹烯,说明T2代转Tafps-B1拟南芥挥发物中含有β-石竹烯。
从上述结果看出,与野生型拟南芥相比,T2代转Tafps-B1拟南芥产生的挥发物中表达量出现明显变化并属于倍半萜的峰为出现在35.889min的峰为β-石竹烯(图5D和E)。
根据人工合成的100ng/μlβ-石竹烯标准样品在GC结果的峰面积为788.8,利用公式((100/788.8)×测试样品峰面积×500)/6株/3d计算野生型拟南芥、T2代转空载体拟南芥和T2代转Tafps-B1拟南芥每棵植株每天挥发物的β-石竹烯的释放量,野生型拟南芥为23.59ng,T2代转Tafps-B1拟南芥的释放量为130.30ng,是野生型拟南芥释放量的5.5倍。T2代转空载体拟南芥和野生型拟南芥结果无显著差异。
三、Tafps-B1在驱避蚜虫中的应用
采用上述二的2的A的方法进行拟南芥气体收集,拟南芥为上述花期(播种后第56天)T2代转Tafps-B1拟南芥、花期野生型拟南芥;分别得到野生型拟南芥挥发物样品(花期)和T2代转Tafps-B1拟南芥挥发物样品(花期)。
用4通道嗅觉仪检测上述花期各植株挥发物对有翅桃蚜对的取食反应的影响,具体如下:
培养室捕获被测试桃蚜虫,每个培养皿中只放一只蚜虫,将其转移到被测屋子中后放置15min以上,以让其适应周围环境。4通道嗅觉仪由3层6mm的有机玻璃的平板组成,塑料螺丝和螺母固定将三层平板固定。最底层的平板上铺一层滤纸方便蚜虫行走。真空泵将嗅觉仪中的气体从中间抽走(流速200ml/min),4个玻璃臂细头用纱布包裹后插入嗅觉仪上。10μl样品滴在滤纸上后放入玻璃臂内。每2min将嗅觉仪顺时针旋转90°,一共持续检测16min。每个样品需要检测10只蚜虫以上。单因素实验测试时,处理臂1放检测T2代转Tafps-B1拟南芥挥发物样品(花期),处理臂2、3、4放溶剂乙醚;双因素选择实验测试时,处理臂1放野生型拟南芥挥发物样品(花期),处理臂3放T2代转Tafps-B1拟南芥挥发物样品,处理臂2、4放溶剂乙醚。实验完毕后,分别用洗洁精、80%酒精和超纯水清洗嗅觉仪,自然晾干;分别用洗洁精、丙酮和超纯水清洗玻璃臂,150℃过夜烘干。所记录得数据利用Excel求平均值及标准差,利用GenStar软件进行显著性分析。
结果如图6所示,为4通道嗅觉仪检测花期转Tafps-B1的拟南芥挥发物对有翅桃蚜对的取食反应的影响,其中(A)为蚜虫分别在野生型拟南芥样品与溶剂乙醚通道的停留时间对比;(B)为蚜虫分别在T2代转Tafps-B1拟南芥挥发物样品与溶剂乙醚的停留时间对比。(C)蚜虫分别在野生型拟南芥挥发物样品、T2代转Tafps-B1拟南芥挥发物样品以及溶剂乙醚的停留时间对比。Tafps1:T2代转Tafps-B1拟南芥挥发物样品;wt:野生型拟南芥挥发物样品;control:溶剂(乙醚)。误差线代表标准误;图B中a表示经t测验转基因株系与对照溶剂差异极其显著(P<0.01,df=10);b表示经t测验对照溶剂与转基因株系差异极其显著(P<0.01,df=10);图C中a表示经t测验转基因株系与野生型b差异显著(P<0.05,df=6);b表示经t测验野生型株系与转基因株系a差异显著(P<0.05,df=6);ab表示经t测验转基因株系和野生型株系与溶剂的差异都不显著;
从图中看出,在单因素实验中,对照为溶剂乙醚,处理通道为挥发物样品。在野生型拟南芥挥发物样品(花期)和乙醚的实验中,蚜虫在野生型拟南芥挥发物样品(花期)通道中停留时间为(3.60±0.72)min,在乙醚通道停留的平均时间为(3.24±0.28)min(图6A);在T2代转Tafps-B1拟南芥挥发物样品(花期)和乙醚的实验中,蚜虫在T2代转Tafps-B1拟南芥挥发物样品(花期)通道中停留时间为(2.55±0.33)min,在乙醚通道停留的平均时间为(3.96±0.13)min(图6B)。通过停留时间的比较计算,表明野生型对有翅桃蚜具有微弱的吸引作用,T2代转Tafps-B1拟南芥挥发物样品对蚜虫都具有显著的驱避作用(P<0.01,df=10)。
利用转基因株系和野生型株系两因素选择实验对有翅桃蚜的行为反应进行生物测试,在野生型拟南芥挥发物样品驱避、T2代转Tafps-B1拟南芥挥发物样品驱避和乙醚的实验中,蚜虫在野生型拟南芥挥发物品通道中停留时间为(5.08±0.69)min,在T2代转Tafps-B1拟南芥挥发物样品通道中停留时间为(2.07±0.47)min,在乙醚通道停留的平均时间为(3.35±0.20)min。通过停留时间的计算,表明与野生型拟南芥相比,T2代转Tafps-B1拟南芥株系具有显著的驱避作用(P<0.05,df=6),见图6C。
采用同样的方法检测花期T2代转空载体拟南芥挥发物,结果与野生型物显著差异。
因此,利用花期所收集的挥发物对对有翅桃蚜对的取食反应行为进行检测。
一种与倍半萜合成相关的蛋白及其编码基因与应用专利购买费用说明
![]()
Q:办理专利转让的流程及所需资料
A:专利权人变更需要办理著录项目变更手续,有代理机构的,变更手续应当由代理机构办理。
1:专利变更应当使用专利局统一制作的“著录项目变更申报书”提出。
2:按规定缴纳著录项目变更手续费。
3:同时提交相关证明文件原件。
4:专利权转移的,变更后的专利权人委托新专利代理机构的,应当提交变更后的全体专利申请人签字或者盖章的委托书。
Q:专利著录项目变更费用如何缴交
A:(1)直接到国家知识产权局受理大厅收费窗口缴纳,(2)通过代办处缴纳,(3)通过邮局或者银行汇款,更多缴纳方式
Q:专利转让变更,多久能出结果
A:著录项目变更请求书递交后,一般1-2个月左右就会收到通知,国家知识产权局会下达《转让手续合格通知书》。











动态评分
0.0