

IPC分类号 : C12N1/21,C12N1/19,C12N1/15,C12N15/63,C12N15/52,C12P7/00,C12P13/00




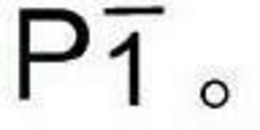
专利摘要
本发明提供了代谢改良的微生物,其特性在于相较野生型生物体有增加的酮酸流,并且包含至少一个编码酶的多核苷酸,相较野生型生物体在表达时引起更高量的化学产物生成。所述重组微生物用于从各种含氮生物质组合物和其他碳源生产大量化学组合物。更特异地,本发明提供了从合适的富氮生物质中生产下列物质的方法:醇、乙醛、乙酸、异丁醛、异丁酸、正丁醛、正丁酸、2-甲基-1-丁醛、2-甲基-1-丁酸、3-甲基-1-丁醛、3-甲基-1-丁酸、氨、铵、氨基酸、2,3-丁二醇、1,4-丁二醇、2-甲基-1,4-丁二醇、2-甲基-1,4-丁二胺、异丁烯、衣康酸盐、乙偶姻、丙酮、异丁烯、1,5-二氨基戊烷、L-乳酸、D-乳酸、莽草酸、甲羟戊酸、聚羟基丁酸(PHB)、类异戊二烯、脂肪酸、高丙氨酸、4-氨基丁酸(GABA)、琥珀酸、苹果酸、柠檬酸、己二酸、对羟基-肉桂酸、四氢呋喃、3-甲基-四氢呋喃、γ-丁内酯、吡咯烷酮、正甲基吡咯烷酮、天冬氨酸、赖氨酸、尸胺、2-酮己二酸、和S-腺苷-甲硫氨酸(SAM)。
权利要求
1.一种重组微生物,其特性在于相较野生型生物体有增加的酮酸流,并且包含至少一个编码酶的多核苷酸,相较野生型生物体在表达时引起更高量的化学产物生成。
2.如权利要求1所述的重组微生物,其特征在于,所述微生物包含编码脱氢酶、转氨酶、和/或脱氨酶的异源多核苷酸。
3.如权利要求2所述的重组微生物,其特征在于,所述脱氢酶选自谷氨酸盐脱氢酶(E.C.1.4.1.2和E.C.1.4.1.4)、谷氨酸脱氢酶(E.C.1.4.1.3)、缬氨酸脱氢酶(E.C.1.4.1.8)、亮氨酸脱氢酶(E.C.1.4.1.9)和苯丙氨酸脱氢酶(E.C.1.4.1.20)。
4.如权利要求3所述的重组微生物,其特征在于,所述亮氨酸脱氢酶是LeuDH。
5.如权利要求4所述的重组微生物,其特征在于,所述亮氨酸脱氢酶LeuDH来自中间型嗜热放线菌。
6.如权利要求2所述的重组微生物,其特征在于,所述脱氨酶选自下组:天冬氨酸解氨酶(4.3.1.1)、L-丝氨酸解氨酶(E.C.4.3.1.17)、D-丝氨酸解氨酶(4.3.1.18)、苏氨酸解氨酶(E.C.4.3.1.19)、酪氨酸解氨酶(E.C.4.3.1.23)、苯丙氨酸解氨酶(E.C.4.3.1.24)、和苯丙氨酸/酪氨酸解氨酶(E.C.4.3.1.25)。
7.如权利要求6所述的重组微生物,其特征在于,所述脱氨酶是丝氨酸脱氨酶SdaB。
8.如权利要求7所述的重组微生物,其特征在于,所述丝氨酸脱氨酶SdaB来自大肠杆菌、玫瑰杆菌、白喉棒状杆菌、肠道沙门氏菌、小肠结肠炎耶尔森菌、或鼻疽伯克霍尔德氏菌。
9.如权利要求2所述的重组微生物,其特征在于,所述转氨酶是L-α-转氨酶(E.C.2.6.1.X,其中X是任意数字)。
10.如权利要求9所述的重组微生物,其特征在于,所述L-α-转氨酶选自L-天冬氨酸转氨酶(E.C.2.6.1.1)、L-丙氨酸转氨酶(E.C.2.6.1.12和E.C.2.6.1.47)、L-天冬酰胺转氨酶(E.C.2.6.1.14)、和甘氨酸转氨酶(E.C.2.6.1.35)。
11.如权利要求10所述的重组微生物,其特征在于,所述L-天冬酰胺转氨酶是AvtA。
12.如权利要求11所述的重组微生物,其特征在于,所述AvtA来自大肠杆菌、脑膜炎奈瑟球菌、菠萝泛菌、地中海拟无枝菌酸菌、曼海姆产琥珀酸菌、肠道沙门菌、或鼠疫耶尔森菌。
13.如权利要求2-12中任一项所述的重组微生物,其特征在于,所述微生物的特征还在于是与野生型生物体相比时降低的氨再摄取活性、降低的2型自诱导物再摄取活性、降低的谷氨酸脱氢酶活性、降低的谷氨酰胺合成酶活性、降低的谷氨酸合成酶活性、降低的群体感应基因活性、和/或降低的整体调节剂活性。
14.如权利要求13所述的重组微生物,其特征在于,所述降低的群体感应活性来自基因luxS或lsrA表达或功能的缺失或降低。
15.如权利要求13所述的重组微生物,其特征在于,所述降低的氨再摄取活性来自基因gdhA或glnA表达或功能的缺失或降低。
16.如权利要求13所述的重组微生物,其特征在于,所述降低的整体调节剂活性来自基因CRP、LRP、Fis和/或IHF表达或功能的缺失或降低。
17.如权利要求1-16中任一项所述的重组微生物,其特征在于,所述化学产物选自醇、乙醛、乙酸、异丁醛、异丁酸、正丁醛、正丁酸、2-甲基-1-丁醛、2-甲基-1-丁酸、3-甲基-1-丁醛、3-甲基-1-丁酸、氨、铵、谷氨酸、苏氨酸、甲硫氨酸、异亮氨酸、缬氨酸、亮氨酸、色氨酸、酪氨酸、苯丙氨酸、2,3-丁二醇、1,4-丁二醇、2-甲基-1,4-丁二醇、2-甲基-1,4-丁二胺、异丁烯、衣康酸盐、乙偶姻、丙酮、异丁烯、1,5-二氨基戊烷、L-乳酸、D-乳酸、莽草酸、甲羟戊酸、聚羟基丁酸(PHB)、类异戊二烯、脂肪酸、高丙氨酸、4-氨基丁酸(GABA)、琥珀酸、苹果酸、柠檬酸、己二酸、对羟基-肉桂酸、四氢呋喃、3-甲基-四氢呋喃、γ-丁内酯、吡咯烷酮、正甲基吡咯烷酮、天冬氨酸、赖氨酸、尸胺、2-酮己二酸、和S-腺苷-甲硫氨酸(SAM)。
18.如权利要求17所述的重组微生物,其特征在于,所述醇选自乙醇、1-丙醇、正丁醇、异丁醇、2-甲基-1-丁醇和3-甲基-丁醇。
19.如权利要求1-16中任一项所述的重组微生物,其特征在于,所述化学产物从丙酮酸生成。
20.如权利要求19所述的重组微生物,其特征在于,所述化学品选自乙醛、乙酸、异丁醛、正丁醛、2,3-丁二醇、L-乳酸、D-乳酸、芳族化合物、聚羟基丁酸(PHB)、甲羟戊酸、类异戊二烯、乙醇、异丁醇、正丁醇、和脂肪酸。
21.如权利要求20所述的重组微生物,其特征在于,所述芳族化合物选自色氨酸、酪氨酸、苯丙氨酸、和莽草酸。
22.如权利要求1-16中任一项所述的重组微生物,其特征在于,所述化学产物从2-酮戊二酸生成。
23.如权利要求22所述的重组微生物,其特征在于,所述化学产物选自GABA(4-氨基丁酸)、谷氨酸、琥珀酸和苹果酸。
24.如权利要求1-16中任一项所述的重组微生物,其特征在于,所述化学品从草酰乙酸盐生成。
25.如权利要求24所述的重组微生物,其特征在于,所述化学品选自天冬氨酸、赖氨酸、尸胺、2-酮己二酸、苏氨酸、甲硫氨酸、和S-腺苷甲硫氨酸(SAM)。
26.如权利要求1-16中任一项所述的重组微生物,其特征在于,所述化学品从2-丁酮酸生成。
27.如权利要求26所述的重组微生物,其特征在于,所述化学品是2-甲基-l-丁醛、异亮氨酸、高丙氨酸、和2-甲基-1-丁醇。
28.如权利要求1-16中任一项所述的重组微生物,其特征在于,所述化学品从2-酮异戊酸生成。
29.如权利要求28所述的重组微生物,其特征在于,所述化学品选自异丁醛、3-甲基-l-丁醛、异丁醇、3-甲基-l-丁醇、和缬氨酸。
30.如权利要求1-16中任一项所述的重组微生物,其特征在于,所述化学品从2-酮异己酸生成。
31.如权利要求30所述的重组微生物,其特征在于,所述化学品选自3-甲基-l-丁醛、亮氨酸、和l-丁醇。
32.如权利要求1-16中任一项所述的重组微生物,其特征在于,所述化学产物从2-酮-3-甲基戊酸生成。
33.如权利要求32所述的重组微生物,其特征在于,所述化学品选自2-甲基-l-丁醛、异亮氨酸、和2-甲基-1-丁醇。
34.如权利要求1-33中任一项所述的重组微生物,其特征在于,所述野生型生物选自细菌、蓝细菌、丝状真菌、和酵母菌。
35.如权利要求34所述的重组微生物,其特征在于,所述野生型生物选自由以下组成的属:梭菌属(Clostridium)、发酵单胞菌属(Zymonomas)、大肠埃希氏菌属(Escherichia)、沙门菌属(Salmonella.)、红球菌属(Rhodococcus)、假单胞菌属(Pseudomonas)、芽孢杆菌属(Bacillus)、乳杆菌属(Lactobacillus)、肠球菌属(Enterococcus)、产碱杆菌属(Alcaligenes)、克雷伯氏菌属(Klesiella)、类芽孢杆菌属(Paenibacillus)、节杆菌属(Arthrobacter)、棒状杆菌属(Corynebacterium)、枯草芽孢杆菌属(Brevibacterium)、毕赤酵母属(Pichia)、念珠菌属(Candida)、汉逊酵母属(Hansenula)、聚球藻属(Synechococcus)、集胞藻属(Synechocystis)、鱼腥藻属(Anabaena)、青枯病(Ralstonia)、乳酸球菌属(Lactococcus)、酵母属(Saccharomyces)、短杆菌属(Brevibacterium)、节细菌属(Arthrobacter)、和微杆菌属(Microbacterium)。
36.如权利要求35所述的重组微生物,其特征在于,所述野生型生物选自大肠杆菌、真养产碱杆菌(Alcaligenes eutrophus)、地衣芽胞杆菌(Bacillus licheniformis)、运动发酵单胞菌(Zymonomas mobilis)、软化芽孢杆菌(Paenibacillus macerans)、红串红球菌(Rhodococcus erythropolis)、恶臭假单胞菌(Pseudomonas putida)、枯草芽孢杆菌(Bacillus subtilis)、胚牙乳杆菌(Lactobacillus plantarum)、谷氨酸棒状杆菌(Corynebacterium glutamicum)、屎肠球菌(Enterococcus faecium)、鹑鸡肠球菌(Enterococcus gallinarium)、粪肠球菌(Enterococcus faecalis)、酿酒酵母(Saccharomyces cerevisiae)、集胞藻(Synechocystis sp.)、鱼腥藻(Anabaena sp.)、真氧产碱杆菌(Ralstonia eutropha)、乳酸乳球菌(Lactococcus lactis)、和细长集球藻(Synechococcus elongatus)。
37.一种通过在有益于生成化学产品的条件下将生物质接触权利要求1-36中任一项所述的重组微生物来从生物质中生产包含蛋白、多肽或氨基酸的化学产物的工艺,其中,生成的化学产品的数量大于野生型生物体生成的数量。
38.如权利要求37所述的工艺,其特征在于,所述生物质选自藻类、干酒糟及其可溶物(DDGS)、细菌、动物残留物、植物、蛋白、多肽、氨基酸或其混合物、和其任意组合。
39.如权利要求38所述的工艺,其特征在于,所述生物质是绿藻、红藻、蓝绿藻、蓝细菌、大肠杆菌、或枯草芽孢杆菌。
40.如权利要求39所述的工艺,其特征在于,所述生物质是小球藻(Chorella vulgaris)、紫球藻(Porphyridium purpureum)、纯顶螺旋藻(Spirulina platensis)、或蓝藻(Synechococcus elongates)。
41.如权利要求37-40中任一项所述的工艺,其特征在于,所述生物质在接触重组微生物前部分降解。
42.如权利要求41所述的工艺,其特征在于,所述生物质用蛋白酶和/或热处理。
43.如权利要求42所述的工艺,其特征在于,所述生物质通过在温度范围60-100℃内加热和通过蛋白酶处理来水解。
44.如权利要求37-43中任一项所述的工艺,其特征在于,所述工艺还包含使所述生物质与所述重组微生物接触、使所述生物质与第二微生物接触,所述第二微生物将赖氨酸、甲硫氨酸、组氨酸、苯丙氨酸、色氨酸和酪氨酸转化成全部20种氨基酸残基的混合物。
45.如权利要求44所述的工艺,其特征在于,所述第二微生物在接触所述重组微生物前与所述生物质接触。
46.如权利要求44或45所述的工艺,其特征在于,所述第二微生物是假单胞菌和/或芽孢杆菌。
说明书
相关申请的交叉引用
本申请要求2010年11月3日提交的美国临时专利申请号61/409,760和2010年11月4日提交的美国临时专利申请序列号61/410,174的优先权,所述申请的公开内容通过引用全文纳入本文。
政府支持
本发明是在美国能源部授予的批准号DE-FC02-02ER63421下由政府资助完成。政府享有本发明的某些权利。
技术领域
提供了代谢修饰的微生物和生成这种微生物的方法。本发明提供了从蛋白质生物质生成生物燃料的生物燃料反应器系统。也提供了通过使合适底物与代谢修饰或重组的微生物和其酶制剂接触生成生物燃料和化学品的方法。
背景技术
因为对经济和环境的关注,预期对生物燃料作为石油替代物的需求增加。所述常见的生物燃料乙醇不是理想的,因为其相比汽油有更低的能量密度,并且必需与汽油在有限的浓度范围内混合以作为运输燃料。乙醇也是吸湿和腐蚀性的,引起存储和分布系统的问题。
另外,己二酸是用于生成包括例如尼龙6-6在内多种合成品的工业目的以及用于制备多种药物的化合物。
再者,γ氨基丁酸是用于生成包括例如尼龙6-6在内多种合成品的工业目的以及生物学作用例如神经细胞通讯的化合物。
发明内容
本公开描述了使用富含氮的材料(如蛋白材料)作为生成多种化合物例如碳燃料和化学品的粗材料。在多个实施方式中,本公开能涉及使用蛋白质作为任何发酵的主要粗材料的方法。所述方法能包含加热和消化细胞生物质以部分破坏蛋白,并且然后使用处理的细胞生物质在多种微生物发酵中作为单一碳源以生成多种生物产品,包含醇、有机酸和其他化学品。在另一个实施方式中,本公开能包含开发特定微生物株以有效利用所述细胞生物质。所述细胞生物质包含所有种类的细菌、酵母、真菌、蓝藻菌、藻类、和任何包含蛋白生物质的粗材料(干酒糟及其可溶物(DDGS)、藻类食物或滤饼、细菌发酵滤饼等)。
在一个实施方式中,开发特定微生物株以具有降解氨基酸的较强能力,从而能利用蛋白生物质(包含蛋白、多肽、氨基酸和其混合物)作为单一碳源。消化多个氨基酸的代谢通路通过遗传和随机突变来工程改造。在某些实施方式中,蛋白和多肽能消化成小肽,其能转运入重组的微生物并且降解成氨基酸。氨基酸转化成多种2-酮酸。最终,这些2-酮酸转化成所需化学实体例如更长链的醛或醇,通过多种重组通路引入到微生物中。其中,亮氨酸、异亮氨酸和缬氨酸直接转化成相应的2-酮酸,这是通过亮氨酸脱氢酶或其他转化这些氨基酸成2-酮酸的酶。丝氨酸、半胱氨酸、色氨酸和丙氨酸及甘氨酸转化成丙酮酸,再转化成2-酮异戊酸,其能再用于例如异丁醇合成。谷氨酸、谷胺酰胺、精氨酸和脯氨酸转化成2-酮戊二酸,再转化成苏氨酸,其能用于例如1-丙醇生成,或能通过苹果酸酶转化回丙酮酸。所述丙酮酸能用于例如异丁醇生成。或者,转化成丙酮酸的所有蛋白生物质也能转化生成乙醇。工程改造残留的氨基酸以支持生长或作为随机突变的能量来源。
在另一个实施方式中,所述蛋白生物质进行处理并转化成多种化学生成的关键中间体,例如丙酮酸、2-酮戊二酸。所述化学品能包含例如但不限于琥珀酸、苹果酸、富马酸和γ-氨基丁酸(GABA)。本发明不限于仅生成所列举的化学品。
本发明在一个实施方式中提供了重组微生物,其特性为相较野生型生物体有增加的酮酸流,并且包含至少一个编码酶的多核苷酸,其相较野生型生物体在表达时生成更高量的化学产物。更特别地,所述重组微生物包含编码脱氢酶、转氨酶、和/或脱氨酶的异源多核苷酸。在某些实施方式中,所述脱氢酶是谷氨酸盐脱氢酶(E.C.1.4.1.2和E.C.1.4.1.4)、谷氨酸脱氢酶(E.C.1.4.1.3)、缬氨酸脱氢酶(E.C.1.4.1.8)、亮氨酸脱氢酶(E.C.1.4.1.9)、或苯丙氨酸脱氢酶(E.C.1.4.1.20)。在常见实施方式中,所述亮氨酸脱氢酶可以是能从中间型嗜热放线菌(Thermoactinomyces intermedius)分离的LeuDH。在某些其他实施方式中,所述重组微生物能包含选自下面的脱氨酶:天冬氨酸解氨酶(4.3.1.1)、L-丝氨酸解氨酶(E.C.4.3.1.17)、D-丝氨酸解氨酶(4.3.1.18)、苏氨酸解氨酶(E.C.4.3.1.19)、酪氨酸解氨酶(E.C.4.3.1.23)、苯丙氨酸解氨酶(E.C.4.3.1.24)、和苯丙氨酸/酪氨酸解氨酶(E.C.4.3.1.25)。更特异地,所述脱氨酶能是丝氨酸脱氨酶SdaB,能获自例如大肠杆菌(Escherichia coli)、玫瑰杆菌(Rosebacter atrosepticum)、白喉棒状杆菌(Corynebacterium diphtheriae)、肠道沙门氏菌(Salmonella enerica)、小肠结肠炎耶尔森菌(Yersinia enterocolitica)、或鼻疽伯克霍尔德氏菌(Burkholderia pseudomallei)。再者,所述重组微生物能包含作为L-α-转氨酶(E.C.2.6.1.X,其中X是任意数字)的转氨酶。在特定实施方式中,所述L-α-转氨酶能是L-天冬氨酸转氨酶(E.C.2.6.1.1)、L-丙氨酸转氨酶(E.C.2.6.1.12和E.C.2.6.1.47)、L-天冬酰胺转氨酶(E.C.2.6.1.14)、或甘氨酸转氨酶(E.C.2.6.1.35)。在某些实施方式中,所述L-天冬氨酸转氨酶能是AvtA,能来自例如大肠杆菌、脑膜炎奈瑟球菌(Neisseria meningitidis)、菠萝泛菌(Pantoea ananatis)、地中海拟无枝菌酸菌(Amycolatopsis mediterranei)、曼海姆产琥珀酸菌(Mannheimia succinicproducens)、肠道沙门菌(Salmonella enterica)或鼠疫耶尔森菌(Yersinia pestis)。
本公开也提供了重组微生物,与野生型生物体相比,其进一步的特性是降低的氨再摄取活性、降低的2型自诱导物再摄取活性、降低的谷氨酸脱氢酶活性、降低的谷氨酰胺合成酶活性、降低的谷氨酸合成酶活性、降低的群体感应基因活性、和/或降低的整体调节剂活性。在某些实施方式中,所述降低的群体感应活性来自基因luxS或lsrA的缺失、或者降低的表达或功能;所述降低的氨再摄取活性来自基因gdhA和glnA的缺失、或者降低的表达或功能,和降低的整体调节剂活性来自基因CRP、LRP、Fis、和/或IHF的缺失、或者降低的表达或功能。
本发明特别公开的重组微生物是生成化学产物例如醇、乙醛、乙酸、异丁醛、异丁酸、正丁醛、正丁酸、2-甲基-1-丁醛、2-甲基-1-丁酸、3-甲基-1-丁醛、3-甲基-1-丁酸、氨、铵、谷氨酸、苏氨酸、甲硫氨酸、异亮氨酸、缬氨酸、亮氨酸、色氨酸、酪氨酸、苯丙氨酸、2,3-丁二醇、1,4-丁二醇、2-甲基-1,4-丁二醇、2-甲基-1,4-丁二胺、异丁烯、衣康酸盐、乙偶姻、丙酮、异丁烯、1,5-二氨基戊烷、L-乳酸、D-乳酸、莽草酸、甲羟戊酸、聚羟基丁酸(PHB)、类异戊二烯、脂肪酸、高丙氨酸、4-氨基丁酸(GABA)、琥珀酸、苹果酸、柠檬酸、己二酸、对羟基-肉桂酸、四氢呋喃、3-甲基-四氢呋喃、γ-丁内酯(butyrolactone)、吡咯烷酮、正甲基吡咯烷酮、天冬氨酸、赖氨酸、尸胺(cadeverine)、2-酮己二酸、和S-腺苷-甲硫氨酸(SAM)。在某些实施方式中,所述醇选自乙醇、1-丙醇(1-proponal)、正丁醇、异丁醇、2-甲基-1-丁醇和3-甲基-丁醇。
再者,本公开提供了所述重组微生物能来自野生型生物体,其是细菌、蓝细菌、丝状真菌、或酵母菌。更特别地,所述野生型微生物来自某一属,例如梭菌属(Clostridium)、发酵单胞菌属(Zymonomas)、大肠埃希氏菌属(Escherichia)、沙门菌属(Salmonella.)、红球菌属(Rhodococcus)、假单胞菌属(Pseudomonas)、芽孢杆菌属(Bacillus)、乳杆菌属(Lactobacillus)、肠球菌属(Enterococcus)、产碱杆菌属(Alcaligenes)、克雷伯氏菌属(Klesiella)、类芽孢杆菌属(Paenibacillus)、节杆菌属(Arthrobacter)、棒状杆菌属(Corynebacterium)、枯草芽孢杆菌属(Brevibacterium)、毕赤酵母属(Pichia)、念珠菌属(Candida)、汉逊酵母属(Hansenula)、聚球藻属(Synechococcus)、集胞藻属(Synechocystis)、鱼腥藻属(Anabaena)、青枯病(Ralstonia)、乳酸球菌属(Lactococcus)、酵母属(Saccharomyces)、短杆菌属(Brevibacterium)、节细菌属(Arthrobacter)、或者微杆菌属(Microbacterium)。更特别地,所述野生型生物体能是例如大肠杆菌、真养产碱杆菌(Alcaligenes eutrophus)、地衣芽胞杆菌(Bacilluslicheniformis)、运动发酵单胞菌(Zymonomas mobilis)、软化芽孢杆菌(Paenibacillus macerans)、红串红球菌(Rhodococcus erythropolis)、恶臭假单胞菌(Pseudomonas putida)、枯草芽孢杆菌(Bacillus subtilis)、胚牙乳杆菌(Lactobacillus plantarum)、谷氨酸棒状杆菌(Corynebacterium glutamicum)、屎肠球菌(Enterococcus faecium)、鹑鸡肠球菌(Enterococcus gallinarium)、粪肠球菌(Enterococcus faecalis)、酿酒酵母(Saccharomyces cerevisiae)、集胞藻(Synechocystis sp.)、鱼腥藻(Anabaena sp.)、真氧产碱杆菌(Ralstonia eutropha)、乳酸乳球菌(Lactococcus lactis)、和细长集球藻(Synechococcus elongatus)。
本公开也提供了重组微生物,所述重组微生物从包含2-酮酸的代谢物和使用氨基酸来源生成所需的化学实体。在一个实施方式中,修饰所述微生物以表达或过量表达有选自亮氨酸或缬氨酸脱氢酶活性的多肽。在另一个实施方式中,工程改造所述微生物以表达或过量表达名为glyA、sda、tnaA、dadX、dadA的酶,其分别用于转化Gly到L-Ser到丙酮酸;L-Cys到丙酮酸;L-到D-Ala到丙酮酸。在另一个实施方式中,所述微生物表达或过量表达选自下列的酶:ybaS、yneH、putA、astABCDE,其分别转化谷胺酰胺、脯氨酸和精氨酸到谷氨酸,然后通过酶gdhA代谢成2-酮戊二酸。
在一个实施方式中,所述重组微生物从工程改造到微生物内的一种或多种多肽表达中生成α-己二酸。在谷氨酸通路中,谷氨酸通过有谷氨酸脱氢酶(gdh)活性的多肽转化成2-KG(2-酮戊二酸)。通过有高柠檬酸合成酶(hcs)活性和高乌头酸酶(homoacornitase)(hacAB)活性的多肽连续反应后,2-KG转化成高异柠檬酸(homoisocitrate),其通过有高异柠檬酸脱氢酶(hicDH)活性的多肽还原脱羧成α酮己二酸。本发明多肽的编码多核苷酸可以来自任意数量的微生物,包括例如酿酒酵母、嗜热栖热菌(Thermus Thermophilus),和本领域其他已知的微生物。所述多核苷酸可以是突变的或变异的酶,来自来源生物体并且然后修饰或工程改造成有提高的活性。在一个实施方式中,所述参与谷氨酸通路的酶的编码多核苷酸从多种生物体中克隆,包含例如酿酒酵母(S.cerevisiae)和嗜热栖热菌(T.Thermophilus)。
在本发明的赖氨酸通路中,赖氨酸通过有赖氨酸转氨酶(lat)活性的多肽脱氨成2-氨基己二酸-6-半醛。然后,有哌啶(piperideine)6-羧酸脱氢酶(pcd)活性的多肽催化形成α-氨基己二酸。所述氨基基团然后通过有2-氨基己二酸氨基转移酶(aadat)活性的多肽脱胺。本发明多肽的编码多核苷酸可以来自任意数量的微生物,包括例如黄杆菌(Flavobacterium lutescens)、棒状链霉菌(Streptomyces clavuligerus)、智人(Homo sapiens)、和本领域其他已知的微生物。所述多核苷酸可以是突变或变异的酶,来自来源生物体并且然后修饰或工程改造成具有提高的活性。在一个实施方式中,所述酶的编码多核苷酸能克隆自多种生物体,包含例如黄杆菌(F.lutescens)、棒状链霉菌、智人等。
对从中间体α酮己二酸生成己二酸而言,工程改造两个生物途径(CoA非依赖性通路和CoA依赖性通路)与一个化学途径,如图3所示。对生物转化而言,α酮己二酸通过各种有脱氢酶活性的多肽转化成α-羟基己二酸,所述多肽包含来自各种微生物的有亮氨酸脱氢酶(ldhA)活性的多肽、有苹果酸脱氢酶(mdh)活性的多肽、和有羟基异己酸(hydroxyisocaproate)脱氢酶(hdh)活性的多肽。在CoA非依赖性通路中,工程改造天然还原性TCA循环的模仿通路,其在厌氧条件下通过苹果酸和富马酸将草酰乙酸盐转化成琥珀酸盐。富马酸还原酶(fumA或fumB)的突变能提供促进α-羟基己二酸脱水的多肽。能通过来自丙酮丁醇梭菌(Clostridium acetobutyricum)的丁烯酸脱氢酶突变体来构建下一步骤。在CoA依赖性通路中,艰难梭菌(Clostridium difficille)中异己酸的生成通路能用于衍生将α羟基己二酸转化成己二酸的多肽。第一步是通过有CoA转移酶(hadA)活性的多肽催化生成R-2-羟基异己酰-CoA。然后,R-2-羟基异己酰-CoA通过脱水酶活化剂复合物(如hadBC hadI)的脱水生成2-异己烯酰(Isocaprenoyl)-CoA,其通过有酰基CoA脱氢酶(acdB etfBA)活性的多肽还原成异己酰-CoA。最终,CoA部分通过CoA转移酶(hadA)除去以生成己二酸。使用包含铂的多种金属催化剂也可能将α酮己二酸化学还原成己二酸。
在一个实施方式中,本公开提供通过引入表达选自下面多肽的异源多核苷酸来修饰的重组细菌:有谷氨酸脱氢酶(gdh)活性的多肽、有高柠檬酸合成酶(hcs)活性的多肽、有高乌头酸酶(hacAB)活性的多肽、有高异柠檬酸脱氢酶(hicDH)活性的多肽、编码前述任意组合的多核苷酸或多种多核苷酸。
在一个实施方式中,本公开提供通过引入表达选自下组多肽的异源多核苷酸来修饰的重组细菌:有赖氨酸转氨酶(lat)活性的多肽、有哌啶6-羧酸脱氢酶(pcd)活性的多肽、有2-氨基己二酸氨基转移酶(aadat)活性的多肽、编码前述任意组合的多核苷酸或多种多核苷酸。
在其他实施方式中,相同或不同的细菌可以重组修饰以表达有脱氢酶活性的多肽,包含来自多种微生物的有亮氨酸脱氢酶(ldhA)活性的多肽、有苹果酸脱氢酶(mdh)活性的多肽、和有羟基异己酸脱氢酶(hdh)活性的多肽。
在另一个实施方式中,本公开提供通过引入表达选自下组多肽的异源多核苷酸来修饰的重组细菌:有CoA转移酶(hadA)活性的多肽、有脱水酶活化剂复合物(如hadBC hadO)活性的多肽、有酰基CoA脱氢酶(acdB etfBA)活性的多肽、有CoA转移酶(hadA)活性的多肽、和任意上述组合,从而从α酮己二酸生成己二酸。
此外,本公开提供另外从包含蛋白、多肽或氨基酸的生物质生成化学产品的工艺,所述工艺通过使生物质在有益于生成化学产品的条件下接触任意上述重组微生物,其中生成的化学产品数量大于野生型生物体生成的数量。所述用于培养重组微生物的生物质能是例如藻类、干酒糟及其可溶物(DDGS)、细菌、动物残留、植物、蛋白、多肽、氨基酸或其混合物、和其任意组合。在本文所述的某些实施方式中,所述生物质能是绿藻、红藻、蓝绿藻、蓝细菌、大肠杆菌或枯草芽孢杆菌。在所述特定实施方式中,所述生物质是小球藻(Chorella vulgaris)、紫球藻(Porphyridium purpureum)、纯顶螺旋藻(Spirulina platensis)、或蓝藻(Synechococcus elongates)。再者,所述生物质能在接触重组微生物前部分降解。在某些实施方式中,所述生物物质能用蛋白酶和/或加热处理。例如,所述生物质通过加热如在温度范围60-100°C内加热,和用蛋白酶处理水解。在某些实施方式中,公开了某一工艺,其中所述生物质能与将赖氨酸、甲硫氨酸、组氨酸、苯丙氨酸、色氨酸和酪氨酸转化成全部20种氨基酸残基混合物的第二微生物接触。特定地,所述第二微生物在接触重组微生物前接触所述生物质。所述第二微生物能是假单胞菌(Pseudomonas)和或芽孢杆菌(Bacillus)。
在附图和以下描述中详细列出了本发明的一个或多个实施方式。通过说明书、附图和权利要求书,可清楚了解其它的特征、目的和优点。
附图简要说明
结合在本说明书中并构成说明书一部分的附图说明了本发明的一个或多个实施方式,并与详细描述一起用来说明本发明的原理和实施。
图1A-E显示了多种生物燃料生产工艺的比较。(A)纤维素植物工艺。(B)藻类脂质工艺。(C)多种生物燃料生产工艺的能量消耗和富N生物质的积累。详细计算见下文。(D)这个研究中开发了藻类蛋白转化成燃料的工艺。(E)来自蛋白的生物燃料的理论产率(产物克数/原材料克数)。实心柱:由排除原材料中的氨基基团计算的净产率。斜纹柱:用原材料中氨基基团计算的总产率。详见表3。图2A-C显示了群体感应失活对生物燃料生产的影响。误差棒表示标准偏差。(A)来自包含不同基因敲除的20个株的相对生物燃料生产效价。野生型株的生物燃料生产效价用作标准化基础。(B和C)YH19(改进氨基酸利用的株)的生物燃料生产(B)和OD(C),及其群体感应基因敲除衍生物,有和没有过量表达所述异丁醇生成通路基因(芽孢杆菌(Bacillus subtilis)alsS、大肠杆菌ilvCD、乳酸乳球菌(Lactococcus lactis)kivd和adhA)。
图3A-E描述了氮中心代谢工程改造策略的整体设计。氨的释放用于驱动氨基酸降解反应以生成2-酮酸。删除gdhA和glnA基因以阻断氨的再摄取。黑色指示过量表达的基因。(A)氨基酸通过直接脱氨和生成NH3来降解。(B)氨基酸通过转氨和生成谷氨酸降解。(C-E)通过IlvE和LeuDH(C),IlvE、AvtA、DadX和DadA(D),和SerC、SerB、SdaB、PpsA、Eno和GpmA(E)设计转氨和脱氨循环。
图4A-D描述了生物燃料生产率的总体设计和优化。(A)开放培养池藻类培养到生物燃料的总体工艺的流程图(600亿加仑(gal)/年,是美国运输燃料消耗的约30%)。在流旁边列举了每个流中C(黑色)、N(灰色)和S(浅灰色)的元素质量流,没有H和O的相关质量。除非另外说明,所有值的单位是百万吨/年。富N生物质意在包含蛋白和糖,尽管使用蛋白到更高级醇的转化率计算。假设藻类物种的脂质含量是10%。正方形盒指示了主要的加工步骤,而椭圆形表示流的成分。每年数十亿加仑表示为Bgal/年。(B)藻类生物质产率作为脂质含量的函数。所述数据(11)从g/L/天转化成g/m2/天,假设培养池深度是0.2m。所述趋势线是图中所示指数函数的最好拟合。(C)考虑到(B)中的生物质生存率和蛋白到生物燃料的理论转化产率,脂质含量对整体生物燃料生产率的影响通过组合的蛋白-脂质工艺(实线)或仅脂质工艺(虚线)来计算。注意到蛋白-脂质工艺的总体生产率随着脂质含量的增加而降低,提示了蛋白部分更有效。(D)组合蛋白-脂质工艺中的最优脂质含量随着作为蛋白到更高级醇转化产率的函数变化。只要蛋白到生物燃料的转化大于理论产率的61%,那么所述工艺的蛋白加工部分比脂质加工更有效。假设从脂质到燃料的碳转化是100%。
图5显示了不同藻类物种、酵母提取物和两个传统蛋白来源之间氨基酸概况的相似性。
图6A-B显示了从各种氨基酸生成更高级醇的碳流驱动策略。(A)碳流驱动更高级醇生成的的代谢网络。氨基酸降解成各种2-酮酸,其通过2-酮酸脱羧酶(KivD)和醇脱氢酶(YqhD)转化成更高级醇。红色指示过量表达的基因。(B)轴下指示了在有过量表达基因的经工程改造大肠杆菌中异丁醇生成的结果。误差棒表示标准偏差。
图7显示了使用氮中心策略工程改造的大肠杆菌中生成生物燃料的链长分布。所述株是YH83(=alsS、ilvC、ilvD、avtA、leuDH、kivD、yqhD、ilvE、ilvA和sdaB的YH19Δg1nAΔLgdhAΔlsrA/过量表达)。
图8显示了使用藻类或细菌细胞水解产物在生长于烧瓶的经工程改造大肠杆菌株YH83中生成的生物燃料(EtOH、iBOH、2MB、3MB)。小的实验室级别的反应器(1升或30升)用于分别培养细菌和藻类细胞。所述藻类生物质混合物包含小球藻(C.vulgaris)、紫球藻(P.purpureum)、纯顶螺旋藻(S.platensis)、或蓝藻(S.elongates)。所有蛋白来源调整为包含21.6g/l的肽和氨基酸,这等同于4%酵母提取物的蛋白量。误差棒表示标准偏差(n=3)。
图9显示了酮酸转化成异丁醇和3-甲基-1-丁醇的总体示意图。
图10显示了酮酸转化成更高级醇的总体示意图。
图11A-C描述了生成己二酸的示意图。
发明详述
本文和所附权利要求书所用的单数形式“一个”、“一种”和“所述”包括复数指示物,除非上下文另有明确说明。因此,例如,提及“一个多核苷酸”包括多个这种多核苷酸,提及“所述微生物”包括一种或多种微生物,等等。
除非另外定义,本文中所使用的所有技术和科学术语都具有本发明所属领域普通技术人员通常所理解的同样含义。虽然在本发明所公开方法和组合物的实施中可以采用类似于或等同于本文所述的那些方法和材料,但是下面描述了示例性的方法、设备和材料。
提供了上面和本文通篇讨论的出版物仅用于其在本申请提交日之前的任何公开。本文中所有内容均不应解释为承认本发明不能凭借在先发明而先于这些公开内容。
本文提供的特定实施例用于显示在生成各种2-酮酸中底物的变化和应用,所述底物包含例如蛋白生物质、蛋白水解物、氨基酸混合物、和单个游离氨基酸等。如下更充分描述,所述2-酮酸然后能进一步代谢成除了生物燃料以外的其他化学实体。
既然植物和藻类不能固定氮,通过植物或藻类的大规模生物燃料生成需要以铵或硝酸盐的形式固定氮,然后主要作为蛋白和少量核酸掺入生物质中。生物氮固定是由一些共生细菌例如根瘤菌和一些独立存活的微生物如鱼腥藻属进行的缓慢和能源密集型过程。非生物氮固定通过已知为哈伯博斯制氨法(Haber Bosch process)的催化方法实现,这也是能源密集型并且有害环境的。生成1摩尔NH3需要8摩尔ATP和4还原当量(通过生物氮固定)或者0.8MJ(通过哈伯博斯制氨法)(57MJ/Kg氮)。
为了生成生物燃料,植物生物质或藻类脂质转化成碳燃料,并且所述剩余富氮生物质用作动物饲料(图1A和1B)。在这些示例中,哈伯博斯制氨法需要固定氮的净输入(图1C)。目前氮肥料的化学合成已经达到每年10000万吨,并且消耗全球初级能量供应的约1.2%,并且如果所述生物燃料工艺按比例增加,所述合成会显著(substantially)增加。用作动物饲料的富氮残留物通过动物废料直接生成温室气体(GHG)。尽管动物饲料现在要求更高的价格,市场不久就会饱和(图1C)。美国畜牧业对富氮生物燃料残留物(即干酒糟及其可溶物,DDGS)的全部潜在需求是约4千2百万吨,对应植物燃料的154亿加仑副产品或者藻类生物燃料的43亿加仑副产品,分别是美国总共液体燃料使用的约7.7%或2.1%。因此,没有氮再循环时,超过上面水平的大规模生物燃料生产会造成地球上还原的氮或GHG的净积累,并且所述实践变成不可持续。
生物燃料生成的两个主要方案之间(图1A和1B),藻类通常比植物长得更快(倍增时间小于24小时),并且不与食物生成竞争。然而,藻类生物质包含大量蛋白(图5),这需要较大净氮输入。另外,快速生长物种包含更高量的蛋白和更低量的液体。通常,氮限制用于以生长速率为代价而提高脂质积累。因此,所述脂质生成物种必需在封闭生物反应器中培养,或者面对被富蛋白天然物种占据的可能性。这种情况产生悖论:或者使用昂贵的封闭培养系统,或者使用有低脂质含量的物种。为了解决这个问题,在氮再循环的开放培养池中使用天然、快速生长藻类物种是理想的,因为这些物种适应局部盐度、温度和pH。既然所述快速生长物种通常含高蛋白和低脂质,就燃料生成和固定氮再循环使用富蛋白生物质能显著降低氨合成的能量成本和间接GHG排放(图1D),而享用低成本开放培养系统。
不像与木质素形成顽固复合物的纤维素,单细胞蛋白能容易被蛋白酶水解成短肽和氨基酸,其已经有各种工业应用。肽键解离能(308KJ/mol)低于纤维素中的β-糖苷键(360KJ/mol),并且更容易断裂。另外,不需要完全蛋白水解,因为短肽能被微生物利用。蛋白酶(量级为100-1000s-1的Kcat是比纤维素酶(甚至就预处理后的可溶底物而言,Kcat是0.1-10s-1量级)更有效的酶,并且很多微生物能天然分泌蛋白酶,提供了一个步骤中最终固结的生物加工的可能性。如下更充分描述,本发明显示了包含微生物蛋白生物质的底物能易于被预处理和酶促水解的简单过程水解。作为示例,预处理能由水中煮沸10分钟组成。酶促水解能通过37°C最小加载蛋白酶4小时完成。这种蛋白生物质水解工艺不需要任何高压、高温、化学加载、更长的预处理、或更长的酶促水解时间,其在纤维素水解工艺中是常见的。
为了将作为底物的肽和氨基酸转化成生物燃料而同时再循环氮,能通过酶如脱氨酶、氨基转移酶或脱氢酶催化的脱胺反应从氨基酸剪切氨基以形成铵。这些反应的很多产物是2-酮酸,其能转化成多种化学产品。其他氨基酸降解成TCA循环中间体,能通过葡萄糖异生作用酶如苹果酸酶或磷酸烯醇丙酮酸羧激酶定向成丙酮酸,并且然后通过乙酰羟酸合成酶或异丙基苹果酸合成酶链延长通路转化成更长酮酸。
所有氨基酸能通过多种生物体组装的通路全部降解,并且所述单个氨基酸到醇(C2-C5)的最大理论产率是35-84%(表1A)。如果蛋白而不是所述水解产物用作计算基础,或者如果在计算净产率中排除所述氨基,所述产率甚至会更高(表1B和C)。通过使用常见藻类氨基酸组合物(图5),来自藻类蛋白的组合醇的最大理论产率计算为60%(图1E;表2A和B)或73%(净产率,计算中排除氮),高于来自糖的乙醇的产率(50%)。]
不幸的是,肽或氨基酸是较差的发酵底物,因为微生物偏好使用氨基酸用于生长而不是形成产物。氨基酸为微生物提供碳和氮。类似地,多种调节系统在适当位置以确保就细胞或微生物群落益处而言的最优营养物消耗。把氨基酸转化成更高级醇的天然通路仅支持少量产物形成。这种低效很大程度归因于不利的热力学梯度,所述梯度驱动转氨反应趋向氨基酸生物合成方向。因此,细菌中所述氨基酸代谢网络需要工程改造。
表1A。来自单个氨基酸的单个生物燃料的理论最大产率(每克氨基酸的醇克数)。来自游离氨基酸的总产率。
表1B。来自单个氨基酸的单个生物燃料的理论最大产率(每克氨基酸的醇克数)。来自游离氨基酸的总产率。一分子肽基氨基酸获得一分子水以形成水解中的游离氨基酸。
表1C来自单个氨基酸的单个生物燃料的理论最大产率(每克氨基酸的醇克数)。来自肽基氨基酸的净产率。在计算净产率时,因为氨要再循环,所述氨分子的分子量从各肽基氨基酸中推断得到。
表2A。来自常见微生物质量的理论最优产物分布。小球藻的氨基酸概况用作输入质量流以计算达到最大醇生成时单个产物的质量生成。
表2B。来自不同蛋白来源的理论生物燃料质量产率。在最右边一列,所述14aa指示了能转化成生物燃料经工程改造大肠杆菌的氨基酸。
本公开提供了利用本公开重组微生物的化学生成系统,作为整体化学生成系统的一部分。例如,本公开提供了图4所示的生物燃料生成系统,其中在开放培养池中日光下产生藻类以生成生物质。利用所述生物质的脂质含量以生成生物柴油,而水解富氮生物质作为本公开重组微生物的底物。所述水解的富氮生物质然后加入包含本公开重组微生物的反应器系统。所述重组微生物代谢富氮生物质以生成多种化学品(依赖于特定重组通路,例如生成醇(例如更高级醇、己二酸等)的酶通路)。所述铵、CO2和副产品然后加回在开放培养池中生长的藻类以支持藻类生长。残留蛋白(如残留的富N生物质)然后能加入第二生物反应器系统以进一步再循环多种代谢物。图4的氮和碳中性生物燃料工艺的示意图利用日光作为能量来源,并且所述氮源是CO2。所述氮和硫来源是工艺中循环的氨和硫酸盐。频繁或持续收获条件下的天然选择通常会偏好快速生长的和稳健的生物体。收获后,首先加工所生成的生物质以就传统生物柴油生成获得脂质。水解蛋白部分,并且加入到第一阶段转换器以生成更高级醇。来自第一阶段的残留氨基酸然后加入第二阶段以生成循环返回所述水解单位的其他生物质。在两阶段转化中生成的CO2能返回藻类培养以富集获自大气的CO2。
在系统的一个实施方式中,所述藻类是富含蛋白或任何其他富含蛋白的光合自养薇生物的藻类。
这个设计的好处是其利用快速生长藻类物种和富N生长条件,促进高蛋白的生长和低脂质的积累(图4B)。理论衍生也显示了高蛋白产率物种有更高的生长速率。确实,所述高蛋白物种的生物质产率比高脂质物种高约10倍(图4B),主要是因为蛋白呈指数式复制,而脂质是线性增加。使用藻类生物质产率和脂质含量之间的关系以及脂质和蛋白转化的理论燃料产率,能计算生物燃料的整体产率(图4C)。如果仅仅利用藻类生物质的脂质部分,随着脂质含量从0增加到20%,整体产率增加,但是当脂质含量增加超过20%时则下降(图4C,虚线)。当认为藻类物种的脂质含量增加实际上并不增加整体燃料产率时,此结果恒定。如果使用藻类生物质的脂质和蛋白部分,然后随着脂质含量增加,所述整体产率实际降低,指示了所述蛋白处理相比脂质处理在整体产率方面更有效。
本公开提供了经代谢工程改造、或重组的微生物,所述微生物包含从合适底物生成各种化合物的生物化学通路,所述底物例如包含单个氨基酸残基的底物、氨基酸残基混合物以及富N生物质。本发明的经代谢工程改造微生物包含生物体基因组内或生物体基因组外部的一个或多个重组多核苷酸。所述微生物包含编码脱氢酶、脱氨酶和/或转氨酶的多核苷酸序列。
所述微生物的特性还能是与野生型生物体相比时,有降低的氨再摄取活性,降低或破坏的群体感应系统例如降低或破坏的2型自诱导物(AI-2)的再摄取,降低的整体调节剂活性。这种降低能通过降低破坏或阻断与氨再摄取、群体感应例如2型自诱导物、和/或整体调节剂活性有关的基因完成。同样,所述微生物能包含选自下组的基因降低、破坏或敲除:在野生型生物体内发现的glnA、gdhA、lsrA、luxS、CRP、LRP、Fis、和/或IHF基因或其任意组合,并且也能包含引入异源多核苷酸。
在一个实施方式中,本公开提供了重组微生物,所述微生物包含与母体微生物相比至少一个靶标酶表达的增加,或者所述微生物编码母体生物体中未发现的酶。所述微生物能包含参与下列的多肽的至少一个编码基因的下降、破坏或敲除:氨再摄取、2型自诱导物(AI-2)再摄取、整体调节剂、谷氨酸脱氢酶、谷氨酰胺合成酶、和/或谷氨酸合成酶,所述基因选自glnA、gdhA、lsrA、luxS、CRP、Lrp、Fis、IHF或其任意组合。
上述实施方式的重组微生物生成增加的酮酸流,并且因此生成参与生物合成通路的至少一个代谢物,所述生物合成与野生型微生物相比时生成化合物的水平增加。所述化合物能是例如但不限于醇、乙醛、乙酸、异丁醛、异丁酸、正丁醛、正丁酸、2-甲基-1-丁醛、2-甲基-1-丁酸、3-甲基-1-丁醛、3-甲基-1-丁酸、氨、铵、谷氨酸、苏氨酸、甲硫氨酸、异亮氨酸、缬氨酸、亮氨酸、色氨酸、酪氨酸、苯丙氨酸、2,3-丁二醇、1,4-丁二醇、2-甲基-1,4-丁二醇、2-甲基-1,4-丁二胺、异丁烯、衣康酸盐、乙偶姻、丙酮、异丁烯、1,5-二氨基戊烷、L-乳酸、D-乳酸、莽草酸、甲羟戊酸、聚羟基丁酸(PHB)、类异戊二烯、脂肪酸、高丙氨酸、4-氨基丁酸(GABA)、琥珀酸、苹果酸、柠檬酸、己二酸、对羟基-肉桂酸、四氢呋喃、3-甲基-四氢呋喃、γ-丁内酯、吡咯烷酮、正甲基吡咯烷酮、天冬氨酸、赖氨酸、尸胺、2-酮己二酸、和S-腺苷-甲硫氨酸(SAM)等。所述醇能是来自合适底物的更高级醇,包含例如异丁醇、1-丁醇、1-丙醇、2-甲基-l-丁醇、3-甲基-l-丁醇和2-苯乙醇。通常,所述重组微生物包含至少一个重组代谢通路,所述通路包含靶标酶和可选还能包含选自下组酶的活性或表达下降:glnA、gdhA、lsrA、luxS、CRP、LRP、Fis、和/或IHF或其任意组合。所述通路用于修饰在生成上面所提供化学组合物中的底物或代谢中间体。所述靶标酶由衍生自合适生物来源的多核苷酸编码或表达。在一些实施方式中,所述多核苷酸包含来自细菌或酵母来源和重组工程改造到本公开微生物中的基因。
本文使用的术语"经代谢工程改造的"或"代谢工程改造"涉及生物合成基因、操纵子相关基因、和这种多核苷酸控制元件的合理通路设计和组装,以用于生成所需代谢物如2-酮酸中增加的流。微生物中,能设计其他通路且生物合成基因、操纵子相关基因和这种多核苷酸的控制元件组装用于在微生物中增加生成和从头生成多种化合物,所述化合物包含例如醇、乙醛、乙酸、异丁醛、异丁酸、正丁醛、正丁酸、2-甲基-1-丁醛、2-甲基-1-丁酸、3-甲基-1-丁醛、3-甲基-1-丁酸、氨、铵、谷氨酸、苏氨酸、甲硫氨酸、异亮氨酸、缬氨酸、亮氨酸、色氨酸、酪氨酸、苯丙氨酸、2,3-丁二醇、1,4-丁二醇、2-甲基-1,4-丁二醇、2-甲基-1,4-丁二胺、异丁烯、衣康酸盐、乙偶姻、丙酮、异丁烯、1,5-二氨基戊烷、L-乳酸、D-乳酸、莽草酸、甲羟戊酸、聚羟基丁酸(PHB)、类异戊二烯、脂肪酸、高丙氨酸、4-氨基丁酸(GABA)、琥珀酸、苹果酸、柠檬酸、己二酸、对羟基-肉桂酸、四氢呋喃、3-甲基-四氢呋喃、γ-丁内酯、吡咯烷酮、正甲基吡咯烷酮、天冬氨酸、赖氨酸、尸胺、2-酮己二酸、和S-腺苷-甲硫氨酸(SAM)等。"经代谢工程改造的"还能包含通过调节和优化转录、翻译、蛋白稳定性和蛋白功能来优化代谢流,使用基因工程改造和合适的培养条件,包含降低、破坏或敲除与引起所需通路的中间体竞争的竞争性代谢通路。生物合成基因能对宿主微生物异源,这或是通过对宿主外源,或是通过突变、重组、和/或与内源宿主细胞中的异源表达控制序列相关联来修饰。一方面,当所述多核苷酸对宿主生物体是自然发生时,所述多核苷酸可以是密码子优化的。
术语"生物合成通路"也称为"代谢通路",指转化(转变)一种化学物质到另一种的一组合成代谢或分解代谢的生物化学反应。如果其平行或顺序作用于相同的底物、生成相同的产物、或作用于或生成相同底物和代谢终产物之间的代谢中间体(即代谢物),那么基因产物属于相同的"代谢通路"。
例如,所述leuABCD操纵子包含leuA、leuB、leuC和leuD基因。其中,leuA编码α-异丙基苹果酸合成酶,leuB编码β-异丙基苹果酸脱氢酶,leuC和leuD编码α-异丙基苹果酸异构酶。这些酶中,α-异丙基苹果酸合成酶催化从α-酮异戊酸到α-异丙基苹果酸的合成反应,α-异丙基苹果酸异构酶催化从α-异丙基苹果酸到β-异丙基苹果酸的异构化反应,和β-异丙基苹果酸脱氢酶催化从β-异丙基苹果酸到α-酮异己酸的脱氢反应,α-酮异己酸是L-亮氨酸生物合成的最终中间体。埃希氏菌(Escherichia)有四种转氨酶,即aspC基因编码的转氨酶A(天冬氨酸谷氨酸氨基转移酶),由ilvGMEDA操纵子所含ilvE基因编码的转氨酶B(BCAA氨基转移酶),avtA基因编码的转氨酶C(丙氨酸缬氨酸氨基转移酶)和tyrB基因编码的转氨酶D(酪氨酸氨基转移酶)。这些酶参与多种氨化反应。这些酶中,转氨酶B和转氨酶D催化上述从α-酮异己酸到L-亮氨酸的氨化反应,反之亦然。转氨酶C和转氨酶D催化L-缬氨酸生物合成通路的最终步骤,所述通路包含L-缬氨酸生物合成和L-亮氨酸生物合成之间的共有通路。
同样,leuABCD操纵子的表达被L-亮氨酸抑制。编码乙酰羟酸合酶I的ilvBN基因的表达受到L-缬氨酸和L-亮氨酸的协同抑制,编码乙酰羟酸合酶II的ilvGM基因的表达受到L-异亮氨酸、L-缬氨酸和L-亮氨酸的协同抑制,和编码乙酰羟酸合酶III的ilvIH基因的表达受到L-亮氨酸的抑制。
术语"底物"或"合适底物"指通过酶作用转化成或意味着要转化成另一化合物的任何物质或化合物。所述术语不仅包含单个化合物,也包含化合物的组合,例如包含至少一个底物或其衍生物的溶液、混合物和其他材料。另外,所述术语"底物"不仅包含提供适用作起始材料的碳源的化合物,例如来自糖或CO2的任意生物质,而且也包含用于本文所述经代谢工程改造微生物相关通路的中间体和终产物代谢物。"生物质衍生糖"包括但不限于分子例如葡萄糖、蔗糖、甘露糖、木糖和阿拉伯糖。术语"生物质衍生糖"包含微生物通常使用的合适碳底物,例如6碳糖,包括但不限于葡萄糖、乳糖、山梨糖、果糖、艾杜糖、半乳糖和甘露糖,所有都是D或L型;或6碳糖组合,例如葡萄糖和果糖;和/或6碳糖酸,包括但不限于2-酮-L-古洛糖酸、艾杜糖酸(IA)、葡糖酸(GA)、6-磷酸葡萄糖酸、2-酮-D-葡糖酸(2-KDG)、5-酮-D-葡糖酸、2-酮葡萄糖磷酸、2,5-二酮-L-古洛糖酸、2,3-L-二酮古洛糖酸、脱氢抗坏血酸、异抗坏血酸(EA)和D-甘露糖酸。富氮生物质指在能水解和纳入氮的例如蛋白、肽、氨基酸、氨基酸混合物、核酸或其他分子富集的生物质。这种富氮生物质包含大蛋白以及更小的肽(如二、三或更长的肽)、氨基酸和氨基酸混合物。
术语"醇"包含例如1-丙醇、异丁醇、1-丁醇、2-甲基-1-丁醇、3-甲基-1-丁醇或2-苯乙醇。
本文提供的重组微生物能表达多种靶标酶,所述酶参与使用合适的碳底物和/或氮源生成例如醇、乙醛、乙酸、异丁醛、异丁酸、正丁醛、正丁酸、2-甲基-1-丁醛、2-甲基-1-丁酸、3-甲基-1-丁醛、3-甲基-1-丁酸、氨、铵、谷氨酸、苏氨酸、甲硫氨酸、异亮氨酸、缬氨酸、亮氨酸、色氨酸、酪氨酸、苯丙氨酸、2,3-丁二醇、1,4-丁二醇、2-甲基-1,4-丁二醇、2-甲基-1,4-丁二胺、异丁烯、衣康酸盐、乙偶姻、丙酮、异丁烯、1,5-二氨基戊烷、L-乳酸、D-乳酸、莽草酸、甲羟戊酸、聚羟基丁酸(PHB)、类异戊二烯、脂肪酸、高丙氨酸、4-氨基丁酸(GABA)、琥珀酸、苹果酸、柠檬酸、己二酸、对羟基-肉桂酸、四氢呋喃、3-甲基-四氢呋喃、γ-丁内酯、吡咯烷酮、正甲基吡咯烷酮、天冬氨酸、赖氨酸、尸胺、2-酮己二酸、和S-腺苷-甲硫氨酸(SAM)等。在特定实施方式中,所述重组微生物包含选自glnA、gdhA、lsrA、luxS或其任意组合的基因或同源物的敲除。这种敲除使微生物的代谢流从氨基酸积累移向生成更多量酮酸、增加的酮酸流、并且然后生成所需上述化合物。
因此,代谢"工程改造"、"修饰"、或"重组"微生物通过将遗传材料引入到宿主或所选母体微生物来生成,因而修饰或改变微生物的细胞生理学和生物化学。通过引入遗传材料,所述母体微生物获得新的属性,如能生成新的、更高量的胞内代谢物。在示例性实施方式中,引入遗传材料到母体微生物产生新的或改良能力以生成所需化合物,这是通过代谢或由富氮生物质到2-酮酸,并且最终到例如醇、乙醛、乙酸、异丁醛、异丁酸、正丁醛、正丁酸、2-甲基-1-丁醛、2-甲基-1-丁酸、3-甲基-1-丁醛、3-甲基-1-丁酸、氨、铵、谷氨酸、苏氨酸、甲硫氨酸、异亮氨酸、缬氨酸、亮氨酸、色氨酸、酪氨酸、苯丙氨酸、2,3-丁二醇、1,4-丁二醇、2-甲基-1,4-丁二醇、2-甲基-1,4-丁二胺、异丁烯、衣康酸盐、乙偶姻、丙酮、异丁烯、1,5-二氨基戊烷、L-乳酸、D-乳酸、莽草酸、甲羟戊酸、聚羟基丁酸(PHB)、类异戊二烯、脂肪酸、高丙氨酸、4-氨基丁酸(GABA)、琥珀酸、苹果酸、柠檬酸、己二酸、对羟基-肉桂酸、四氢呋喃、3-甲基-四氢呋喃、γ-丁内酯、吡咯烷酮、正甲基吡咯烷酮、天冬氨酸、赖氨酸、尸胺、2-酮己二酸、和S-腺苷-甲硫氨酸(SAM)等。引入母体微生物的遗传材料包含一种或多种酶的编码基因、或基因部分,所述一种或多种酶参与生成任何上面化合物的生物合成通路,并且也可以包含表达和/或调控这些基因的其他元件,如启动子序列。
如上所述,所述工程改造或修饰的微生物除了引入遗传材料到宿主或母体微生物外,包含破坏、删除或敲除基因或多核苷酸以改变微生物的细胞生理学和生物化学。通过降低、破坏或敲除基因或多核苷酸,所述微生物获得新的或改善的特性(如能生成新的或更多数量的胞内代谢物,提高代谢物例如酮酸的流,下调所需通路,和/或降低不需要副产物的生成)。
例如,本公开显示了有异源kivd、yqhD,或其他醇脱氢酶和ilvA、leuA、leuB、leuC、leuD(或Leu操纵子,如leuABCD)、ansB、aspC、rocG、putA、ybaS、ilvA、glyA、sdaA、tnaA、alsS、ilvCD、ilvE的过量表达或异源表达,能从包含N或蛋白生物质中获得更高级醇的生成。从蛋白材料中生成2-酮酸也能引起从2-酮酸生成感兴趣的其他化学实体,例如醇、乙醛、乙酸、异丁醛、异丁酸、正丁醛、正丁酸、2-甲基-1-丁醛、2-甲基-1-丁酸、3-甲基-1-丁醛、3-甲基-1-丁酸、氨、铵、谷氨酸、苏氨酸、甲硫氨酸、异亮氨酸、缬氨酸、亮氨酸、色氨酸、酪氨酸、苯丙氨酸、2,3-丁二醇、1,4-丁二醇、2-甲基-1,4-丁二醇、2-甲基-1,4-丁二胺、异丁烯、衣康酸盐、乙偶姻、丙酮、异丁烯、1,5-二氨基戊烷、L-乳酸、D-乳酸、莽草酸、甲羟戊酸、聚羟基丁酸(PHB)、类异戊二烯、脂肪酸、高丙氨酸、4-氨基丁酸(GABA)、琥珀酸、苹果酸、柠檬酸、己二酸、对羟基-肉桂酸、四氢呋喃、3-甲基-四氢呋喃、γ-丁内酯、吡咯烷酮、正甲基吡咯烷酮、天冬氨酸、赖氨酸、尸胺、2-酮己二酸、和S-腺苷-甲硫氨酸(SAM)。某些实施方式能包含任何一种或多种上述基因的表达或过量表达,所述一种或多种基因伴随着减少或敲除氨再摄取、群体感应基因活性例如2型自诱导物(AI-2)再摄取、和/或整体调节剂基因等。在特定实施方式中,任何一种或多种上述基因的表达或过量表达也能伴随敲除或降低选自下组的基因或同源物表达:glnA、gdhA、lsrA、luxS、CRP、LRP、Fis、和/或IHF、或其任意组合。生成例如醇、乙醛、乙酸、异丁醛、异丁酸、正丁醛、正丁酸、2-甲基-1-丁醛、2-甲基-1-丁酸、3-甲基-1-丁醛、3-甲基-1-丁酸、氨、铵、谷氨酸、苏氨酸、甲硫氨酸、异亮氨酸、缬氨酸、亮氨酸、色氨酸、酪氨酸、苯丙氨酸、2,3-丁二醇、1,4-丁二醇、2-甲基-1,4-丁二醇、2-甲基-1,4-丁二胺、异丁烯、衣康酸盐、乙偶姻、丙酮、异丁烯、1,5-二氨基戊烷、L-乳酸、D-乳酸、莽草酸、甲羟戊酸、聚羟基丁酸(PHB)、类异戊二烯、脂肪酸、高丙氨酸、4-氨基丁酸(GABA)、琥珀酸、苹果酸、柠檬酸、己二酸、对羟基-肉桂酸、四氢呋喃、3-甲基-四氢呋喃、γ-丁内酯、吡咯烷酮、正甲基吡咯烷酮、天冬氨酸、赖氨酸、尸胺、2-酮己二酸、和S-腺苷-甲硫氨酸(SAM)等涉及多种2-酮酸中间体。
本公开也显示了编码下面多肽的一种或多种异源多核苷酸的表达或一种或多种内源多核苷酸的过量表达用于生成代谢通路以产生任何数目的其他所需化合物。例如,在谷氨酸通路中,谷氨酸通过有谷氨酸脱氢酶(gdh)活性的多肽转化成2-KG(2-酮戊二酸)。通过有高柠檬酸合成酶(hcs)活性和高异乌头酸酶(hacAB)活性的多肽连续反应后,2-KG转化成高异柠檬酸,其通过有高异柠檬酸脱氢酶(hicDH)活性的多肽还原脱羧成α酮己二酸。本发明多肽的编码多核苷酸可以来自任意数量的微生物,包含酿酒酵母、嗜热栖热菌、和本领域其他已知的微生物。所述多核苷酸可以是突变或变异的酶,来自来源生物体并且然后修饰或工程改造成具有提高的表达和/或活性。在一个实施方式中,所述参与谷氨酸通路的酶的编码多核苷酸从多种生物体中克隆,包含酿酒酵母和嗜热栖热菌。
在本发明的赖氨酸通路中,赖氨酸通过有赖氨酸转氨酶(lat)活性的多肽脱氨成2-氨基己二酸-6-半醛。然后,有哌啶6-羧酸脱氢酶(pcd)活性的多肽催化形成α-氨基己二酸。所述氨基团然后通过有2-氨基己二酸氨基转移酶(aadat)活性的多肽脱胺。本发明多肽的编码多核苷酸可以来自任意数量的微生物,包含黄杆菌(F.lutescens)、棒状链霉菌、智人、和本领域其他已知的微生物。所述多核苷酸可以是突变或变异的酶,来自来源生物体并且然后修饰或工程改造成具有提高的表达和/或活性。在一个实施方式中,所述酶的编码多核苷酸能克隆自多种生物体,包含黄杆菌(F.lutescens)、棒状链霉菌、智人、和本领域其他已知的生物体。
例如对从中间体α酮己二酸生成己二酸而言,工程改造两个生物途径(CoA非依赖性通路和CoA依赖性通路)与一个化学途径,如图3所示。对生物转化而言,α酮己二酸转化成α-羟基己二酸,所示转化通过各种有脱氢酶活性的多肽,包含来自各种微生物的有亮氨酸脱氢酶(ldhA)活性的多肽、有苹果酸脱氢酶(mdh)活性的多肽、和有羟基异己酸脱氢酶(hdh)活性的多肽。在CoA非依赖性通路中,工程改造天然还原性TCA循环的模仿通路,其在厌氧条件下通过苹果酸和富马酸将草酰乙酸盐转化成琥珀酸盐。富马酸还原酶基因(例如fumA或fumB)的突变能提供促进α-羟基己二酸脱水的多肽。能通过来自丙酮丁醇梭菌(Clostridium acetobutyricum)的丁烯酸脱氢酶突变来构建下一步骤。在CoA依赖性通路中,艰难梭菌(Clostridium difficille)中异己酸的生成通路能用于衍生将α羟基己二酸转化成己二酸的多肽。第一步是通过有CoA转移酶(hadA)活性的多肽催化生成R-2-羟基异己酰CoA。然后,R-2-羟基异己酰-CoA通过脱水酶活化剂复合物(如hadBC hadI)的脱水生成2-异己烯酰-CoA,其通过有酰基CoA脱氢酶(acdB etfBA)活性的多肽还原成异己酰-CoA。最终,CoA部分通过CoA转移酶(hadA)除去以生成己二酸。使用包含铂的多种金属催化剂也可能将α酮己二酸化学还原成己二酸。在其他实施方式中,所述微生物包含内源谷氨酸生成通路。例如,有谷氨酸生成通路的有用生物体包含来自诸如短杆菌属(Brevibacterium)、节细菌属(Arthrobacter)、微杆菌属(Microbacterium)、棒状杆菌属(Corynebacterium)等属的那些细菌或酵母物种。
有谷氨酸脱氢酶(gdh)的多肽为本领域已知或能重组生产,包含有增加或改善活性的非天然产生的多肽。有谷氨酸脱氢酶活性的示例性多肽能与SEQ ID NO:2所示氨基酸序列(相应核苷酸序列示于SEQ ID NO:1)有80%-99%相同性。例如,谷氨酸脱氢酶能包含来自酿酒酵母或解朊假丝酵母变种(Candida famata var.)的SEQ ID NO:2,有示于GenBank登录号CAQ53143的序列(通过引用纳入本文)。用于本公开方法和组合物的其他合适谷氨酸脱氢酶能使用例如BLAST算法容易鉴定。
有高柠檬酸合成酶的多肽为本领域已知或能重组生产,包含有增加或改善活性的非天然产生的多肽。有高柠檬酸合成酶活性的示例性多肽能与SEQ ID NO:4所示序列(相应核苷酸序列示于SEQ ID NO:3)有80%-99%相同性。例如,高柠檬酸合成酶能包含来自酿酒酵母或嗜热栖热菌的SEQ ID NO:4,有示于GenBank登录号BAA33785的序列(通过引用纳入本文)。用于本公开方法和组合物的其他合适高柠檬酸合成酶能使用例如BLAST算法容易鉴定。
有高乌头酸酶活性的多肽为本领域已知或能重组生产,包含有增加或改善活性的非天然产生的多肽。有高乌头酸酶活性的示例性多肽能与SEQ ID NO:6所示序列(相应核苷酸序列示于SEQ ID NO:5)有80%-99%相同性。例如,高乌头酸酶能包含来自酿酒酵母或皮炎组织胞浆菌(Ajellomyces dermatitidis)的SEQ ID NO:6,有示于GenBank登录号XP_002620204的序列(通过引用纳入本文)。用于本公开方法和组合物的其他合适高柠檬酸合成酶能使用例如BLAST算法容易鉴定。
有高异柠檬酸脱氢酶活性的多肽为本领域已知或能重组生产,包含有增加或改善活性的非天然产生的多肽。有高异柠檬酸脱氢酶活性的示例性多肽能与SEQ ID NO:8所示序列(相应核苷酸序列示于SEQ ID NO:7)有80%-99%相同性。例如,高异柠檬酸脱氢酶多肽能包含来自嗜热栖热菌的SEQ ID NO:8。用于本公开方法和组合物的其他合适高柠檬酸合成酶能使用例如BLAST算法容易鉴定。
有赖氨酸转氨酶活性的多肽为本领域已知或能重组生产,包含有增加或改善活性的非天然产生的多肽。有赖氨酸转氨酶活性的示例性多肽能与SEQ ID NO:10所示序列(相应核苷酸序列示于SEQ ID NO:9)有80%-99%相同性。例如,赖氨酸转氨酶多肽能包含来自棒状链霉菌的SEQ ID NO:10。用于本公开方法和组合物的其他合适赖氨酸转氨酶能使用例如BLAST算法容易鉴定。
有哌啶6-羧酸脱氢酶(pcd)活性的多肽为本领域已知或能重组生产,包含有增加或改善活性的非天然产生的多肽。有哌啶6-羧酸脱氢酶(pcd)活性的示例性多肽能与SEQ ID NO:12所示序列(相应核苷酸序列示于SEQ ID NO:11)有80%-99%相同性。例如,哌啶6-羧酸脱氢酶(pcd)多肽能包含来自黄杆菌(Flavobacterium lutescens)的SEQ ID NO:12。用于本公开方法和组合物的其他合适哌啶6-羧酸脱氢酶(pcd)能使用例如BLAST算法容易鉴定。
有2-氨基己二酸氨基转移酶(aadat)活性的多肽为本领域已知或能重组生产,包含有增加或改善活性的非天然产生的多肽。有2-氨基己二酸氨基转移酶(aadat)活性的示例性多肽能与SEQ ID NO:14所示序列(相应核苷酸序列示于SEQ ID NO:13)有80%-99%相同性。例如,2-氨基己二酸氨基转移酶(aadat)多肽能包含来自智人的SEQ ID NO:14。用于本公开方法和组合物的其他合适2-氨基己二酸氨基转移酶(aadat)使能用例如BLAST算法容易鉴定。
有亮氨酸脱氢酶(ldh)活性的多肽为本领域已知或能重组生产,包含有增加或改善活性的非天然产生的多肽。有亮氨酸脱氢酶活性的示例性多肽能与SEQ ID NO:16所示序列(相应核苷酸序列示于SEQ ID NO:15)有80%-99%相同性。例如,亮氨酸脱氢酶多肽能包含来自嗜热脂肪地芽孢杆菌(Geobacillus stearothermophilus)的SEQ ID NO:16。用于本公开方法和组合物的其他合适亮氨酸脱氢酶能使用例如BLAST算法容易鉴定。
有苹果酸脱氢酶(mdh)活性的多肽为本领域已知或能重组生产,包含有增加或改善活性的非天然产生的多肽。有苹果酸脱氢酶活性的示例性多肽能与SEQ ID NO:18所示序列(相应核苷酸序列示于SEQ ID NO:17)有80%-99%相同性。例如,苹果酸脱氢酶多肽能包含来自大肠杆菌的SEQ ID NO:18。用于本公开方法和组合物的其他合适苹果酸脱氢酶能使用例如BLAST算法容易鉴定。
有羟基异己酸脱氢酶(hdh)活性的多肽为本领域已知或能重组生产,包含有增加或改善活性的非天然产生的多肽。有羟基异己酸脱氢酶活性的示例性多肽能与SEQ ID NO:20所示序列(相应核苷酸序列示于SEQ ID NO:19)有80%-99%相同性。例如,羟基异己酸脱氢酶多肽能包含来自烟曲霉(Aspergillus fumigatus)的SEQ ID NO:20。用于本公开方法和组合物的其他合适羟基异己酸脱氢酶能使用例如BLAST算法容易鉴定。
两个膜结合的含FAD酶负责催化富马酸和琥珀酸的互变;所述富马酸还原酶用于厌氧生长,而所述琥珀酸脱氢酶用于好氧生长。富马酸还原酶包含多个亚基(如大肠杆菌中的frdA、frdB、和frdC)。任意一个亚基的修饰能产生本文所需活性。例如,敲除frdB、frdC或frdBC用于本公开的方法。已知Frd的同源物和变体。例如,同源物和变体包含例如富马酸还原酶亚基D(富马酸还原酶13kDa疏水蛋白)gi|67463543|sp|P0A8Q3.1|FRDD_ECOLI(67463543);富马酸还原酶亚基C(富马酸还原酶15kDa疏水蛋白)gi|1346037|sp|P20923.2|FRDC_PROVU(1346037);富马酸还原酶亚基D(富马酸还原酶13kDa疏水蛋白)gi|120499|sp|P20924.1|FRDD_PROVU(120499);富马酸还原酶亚基C(富马酸还原酶15kDa疏水蛋白)gi|67463538|sp|POA8Q0.1|FRDC_ECOLI(67463538);富马酸还原酶铁硫亚基(大肠杆菌)gi|145264|gb|AAA23438.1|(145264);富马酸还原酶黄素蛋白亚基(大肠杆菌)gi|145263|gb|AAA23437.1|(145263);富马酸还原酶黄素蛋白亚基gi|37538290|sp|P17412.3|FRDA_WOLSU(37538290);富马酸还原酶黄素蛋白亚基gi|120489|sp|P00363.3|FRDA_ECOLI(120489);富马酸还原酶黄素蛋白亚基gi|120490|sp|P20922.1|FRDA_PROVU(120490);富马酸还原酶黄素蛋白亚基前体(Flavocytochrome c)(Flavocytochrome c3)(Fcc3)gi|119370087|sp|Q07WU7.2|FRDA_SHEFN(119370087);富马酸还原酶铁硫亚基gi|81175308|sp|POAC47.2|FRDB_ECOLI(81175308);富马酸还原酶黄素蛋白亚基(Flavocytochrome c)(Flavocytochrome c3)(Fcc3)gi|119370088|sp|POC278.1|FRDA_SHEFR(119370088);Frd操纵子未表征的蛋白C gi|140663|sp|P20927.1|YFRC_PROVU(140663);frd操纵子可能的(probable)铁硫亚基A gi|140661|sp|P20925.1|YFRA_PROVU(140661);富马酸还原酶铁硫亚基gi|120493|sp|P20921.2|FRDB_PROVU(120493);富马酸还原酶黄素蛋白亚基gi|2494617|sp|006913.2|FRDA_HELPY(2494617);富马酸还原酶黄素蛋白亚基前体(铁(III)诱导的flavocytochrome C3)(Ifc3)gi|13878499|sp|Q9Z4P0.1|FRD2_SHEFN(13878499);富马酸还原酶黄素蛋白亚基gi|54041009|sp|P64174.1|FRDA_MYCTU(54041009);富马酸还原酶黄素蛋白亚基gi|54037132|sp|P64175.1|FRDA_MYCBO(54037132);富马酸还原酶黄素蛋白亚基gi|12230114|sp|Q9ZMP0.1|FRDA_HELPJ(12230114);富马酸还原酶黄素蛋白亚基gi|1169737|sp|P44894.1|FRDA_HAEIN(1169737);富马酸还原酶黄素蛋白亚基(产琥珀酸沃廉菌(Wolinella succinogenes))gi|13160058|emb|CAA04214.2|(13160058);富马酸还原酶黄素蛋白亚基前体(flavocytochrome c)(FL cyt)gi|25452947|sp|P83223.2|FRDA_SHEON(25452947);富马酸还原酶铁硫亚基(产琥珀酸沃廉菌)gi|2282000|emb|CAA04215.1|(2282000);和富马酸还原酶细胞色素b亚基(产琥珀酸沃廉菌)gi|2281998|emb|CAA04213.1|(2281998),与登录号有关的各序列通过引用全文纳入本文。
有羟基异己酸CoA转移酶(hadA)活性的多肽为本领域已知或能重组生产,包含有增加或改善活性的非天然产生的多肽。有羟基异己酸CoA转移酶活性的示例性多肽能与SEQ ID NO:22所示序列(相应核苷酸序列示于SEQ ID NO:21)有80%-99%相同性。例如,羟基异己酸CoA转移酶多肽能包含来自艰难梭菌(Clostridium difficile)的SEQ ID NO:22。用于本公开方法和组合物的其他合适羟基异己酸CoA转移酶能使用例如BLAST算法容易鉴定。
有2-羟基异己酰-CoA脱水酶活性的多肽为本领域已知或能重组生产,包含有增加或改善活性的非天然产生的多肽。有2-羟基异己酰-CoA脱水酶活性的示例性多肽与SEQ ID NO:24(对应核苷酸序列示于SEQ ID NO:23)和SEQ ID NO:26(对应核苷酸序列示于SEQ ID NO:25)所示序列能有80%-99%相同性。例如,2-羟基异己酰-CoA脱水酶活性多肽能包含来自艰难梭菌的SEQ ID NO:24或SEQ ID NO:26。用于本公开方法和组合物的其他合适2-羟基异己酰-CoA脱水酶能使用例如BLAST算法容易鉴定。
2-羟基异己酰-CoA激活剂多肽为本领域已知或能重组生产,包含有增加或改善活性的非天然产生的多肽。有2-羟基异己酰-CoA脱水酶活性的示例性多肽能与SEQ ID NO:27所示序列(相应核苷酸序列示于SEQ ID NO:26)有80%-99%相同性。例如,2-羟基异己酰-CoA脱水酶活化多肽能包含来自艰难梭菌(Clostridium difficile)的SEQ ID NO:27。用于本公开方法和组合物的其他合适2-羟基异己酰-CoA脱水酶活化酶能使用例如BLAST算法容易鉴定。
有酰基CoA脱氢酶活性的多肽为本领域已知或能重组生产,包含有增加或改善活性的非天然产生的多肽。有酰基CoA脱氢酶活性的示例性多肽能与SEQ ID NO:29所示序列(相应核苷酸序列示于SEQ ID NO:28)有80%-99%相同性。例如,酰基CoA脱氢酶多肽能包含来自艰难梭菌(Clostridium difficile)的SEQ ID NO:29。用于本公开方法和组合物的其他合适酰基CoA脱氢酶使用例如BLAST算法容易鉴定。
<通过蛋白质生物质发酵的重组微生物的生物燃料和化学生产专利购买费用说明
![]()
Q:办理专利转让的流程及所需资料
A:专利权人变更需要办理著录项目变更手续,有代理机构的,变更手续应当由代理机构办理。
1:专利变更应当使用专利局统一制作的“著录项目变更申报书”提出。
2:按规定缴纳著录项目变更手续费。
3:同时提交相关证明文件原件。
4:专利权转移的,变更后的专利权人委托新专利代理机构的,应当提交变更后的全体专利申请人签字或者盖章的委托书。
Q:专利著录项目变更费用如何缴交
A:(1)直接到国家知识产权局受理大厅收费窗口缴纳,(2)通过代办处缴纳,(3)通过邮局或者银行汇款,更多缴纳方式
Q:专利转让变更,多久能出结果
A:著录项目变更请求书递交后,一般1-2个月左右就会收到通知,国家知识产权局会下达《转让手续合格通知书》。












动态评分
0.0