

IPC分类号 : C12N15/60,C12N9/88,C12N15/11,C12N15/82,C12P5/00,C12Q1/6895,A01H5/00,A01H5/10,A01H6/00





专利摘要
本发明公开了一种姜花倍半萜合成酶基因HcTPS12及其应用。所述HcTPS12基因的全长cDNA序列如SEQIDNO:1所示,编码序列如SEQIDNO:2所示,其编码的氨基酸序列如SEQIDNO:3所示。所述HcTPS12基因在姜花叶组织中表达量较高,在根茎和花等器官几乎不表达,并且其表达量受叶片发育的调控。将HcTPS12的外源重组蛋白催化底物后可以生成倍半萜药用成分红没药烯,可用于制备红没药烯及进一步制备精油、香精和药物;将HcTPS12连接植物转化载体,然后导入姜花或其它植物细胞中,可获得表达所述基因的转基因植株;还根据本发明所述基因序列信息产生特异性的分子标记,用于鉴定姜花或其它植物的倍半萜红没药烯合成酶基因,用于分子标记辅助选择育种,从而提高育种的选择效率,具有较大的应用前景。
权利要求
1.一种姜花倍半萜合成酶基因
2.一种姜花倍半萜合成酶HcTPS12,其特征在于,所述蛋白的氨基酸序列如SEQ ID NO:3所示。
3.一种重组载体,其特征在于,包含权利要求1所述姜花倍半萜合成酶基因
4.一种包含权利要求3所述重组载体的重组菌。
5.一种包含权利要求4所述重组菌的细胞系。
6.权利要求1姜花倍半萜合成酶基因
说明书
技术领域
本发明涉及基因工程技术领域,更具体地,涉及一种姜花倍半萜合成酶基因HcTPS12及其应用。
背景技术
姜花是姜科姜花属多年生草本植物,在印度地区是一种传统的药用植物(Ray,et al.,2016)。从姜花根茎、花瓣和叶片中可以提取出大量精油,研究表明姜花精油具有抗菌、抗氧化及消炎等药用特性,同时广泛应用于护肤品、香精香料等行业,并且主要成分为萜烯类化合物(Santos,et al.,2010;Dixit,2018)。萜类物质是一类由数个异戊二烯(Isoprene,C5)结构单位构成的化合物的统称,根据其结构单位数目的不同,可分为单萜(Monoterpene,C10)、倍半萜(Sesquiterpene,C15)和二萜(Diterpene,C20)等(Martin et al.,2015)。其中倍半萜是重要的药用成分,同时还是重要的香精成分。据报道,倍半萜紫杉醇可以用来治疗肿瘤(吴娜等,2016),倍半萜青蒿素是治疗疟疾的特效药(Mann,et al.,2000)。倍半萜红没药烯具有抗痒、消炎和抗癌等作用,同时是重要食用香精(庞亚如和胡智慧等,2018)。
萜类合成酶(TPS)是萜类物质生物合成的末端关键酶,它直接决定萜类产物的种类、数量和产量。根据TPS催化形成的产物的不同,将TPS分为单萜合成酶(MonoTPS)、倍半萜合成酶(SesquiTPS)和二萜合成酶(DiTPS),它们分别催化底物GPP、FPP和GGPP形成相应的单萜、倍半萜和二萜产物。
萜类合成酶是萜类化合物生物合成的关键酶,因此也成了萜类生物合成过程中研究最多和最深入的酶类,自1992年在烟草中克隆了两个倍半萜合成酶基因(Facchini and Chappell,1992)以来,科学家已在40多种植物中克隆了200多个单萜和倍半萜合成酶基因(Degenhardt et al.,2009),涉及农作物(Chen et al.,1995;Kollner et al.,2008;Yuan et al.,2008;Chen et al.,2014)、针叶类植物(Bohlmann et al.,1999;Martin et al.,2004)、药用植物(Deguerry et al.,2006)、香料植物和观赏植物以及拟南芥(Chen et al.,2003)等模式植物。
目前,国内对姜花萜类合成酶的研究主要集中在单萜合成酶基因上,然而对姜花倍半萜合成酶基因却鲜有报道。克隆倍半萜合成酶基因是对姜花倍半萜成分形成机理研究的前提,为以后全面阐述姜花倍半萜成分形成和调控机理及通过基因工程手段提高姜花倍半萜成分的含量奠定理论基础。
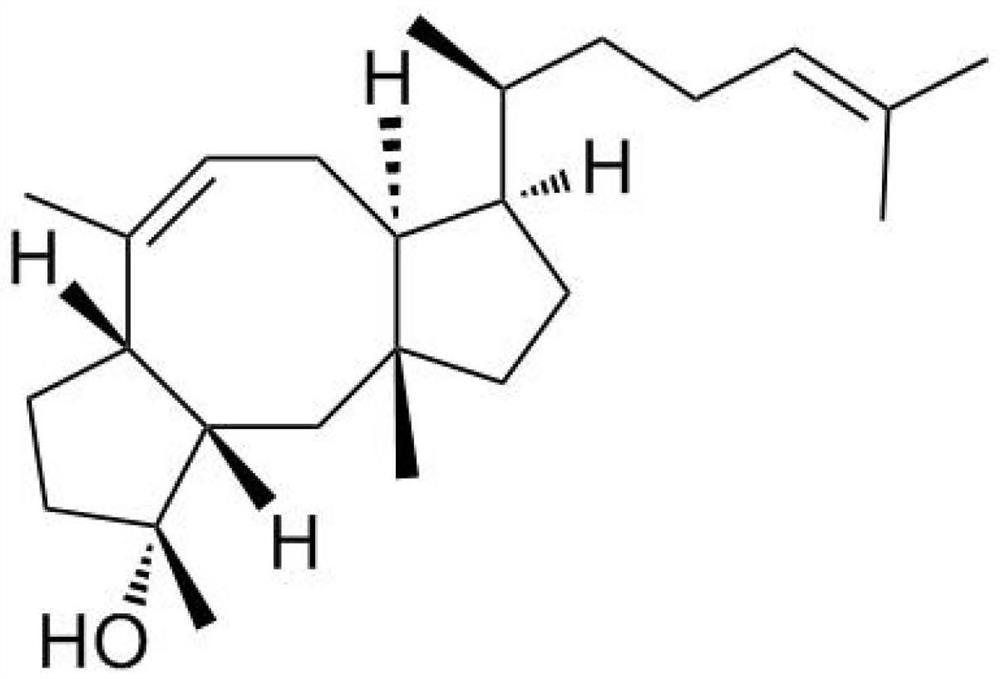
红没药烯是一种植物来源的倍半萜烯天然活性化合物,它在自然界中分布也较为广泛,主要存在于天然的植物精油中。根据双键位置的不同可分为3种同分异构体:α-红没药烯、β-红没药烯和γ-红没药烯;红没药烯有类似果香和香脂的气味,其中β-红没药烯有类似香油的气味,可以作为食用香精及日化香精;红没药烯也是极具潜力的新型生物燃料;β-红没药烯和γ-红没药烯还具有抗痒、消炎的功能和抗癌活性。此外,红没药烯也是多种高附加值工业产品合成的前体物质,如生物燃料、生物塑料、化妆品、保健品及药品等。
目前红没药烯的工业生产主要是通过植物提取法实现的,但从植物组织中提红没药烯存在着产物含量低和分离纯化困难等缺点。微生物代谢工程的快速发展为这些植物天然产物的生产提供了一条更具潜力的生物合成路线。利用微生物代谢工程技术构建生产这些有价值的植物天然产物的微生物细胞工厂具有绿色清洁、可持续发展和经济效益好等独特优势。
发明内容
本发明所要解决的技术问题是克服姜花萜类合成酶基因研究及红没药烯制备方面存在的缺陷和不足,提供一种姜花中控制倍半萜成分红没药烯的单功能酶基因HcTPS12。
本发明的第一个目的是提供一种姜花倍半萜合成酶基因HcTPS12。
本发明的第二个目的是提供一种姜花倍半萜合成酶HcTPS12。
本发明的第三个目的是提供所述姜花倍半萜合成酶基因HcTPS12和/或姜花倍半萜合成酶HcTPS12在制备红没药烯中的应用。
本发明的上述目的是通过以下技术方案给予实现的:
一种姜花倍半萜合成酶基因HcTPS12,所述基因的全长cDNA序列如SEQ ID NO:1所示,所述基因可赋予姜花叶片倍半萜红没药烯成分。本发明所示姜花基因HcTPS12在叶组织中表达量较高,并且其表达与叶发育进程有关。HcTPS12基因cDNA全长为1823bp,其核苷酸序列如SEQ ID NO:1所示;基因编码区(CDS)共1653bp,其核苷酸序列如SEQ ID NO:2所示;推测编码550个氨基酸,其氨基酸序列如SEQ ID NO:3所示,推测蛋白分子量为64.2kDa。该基因序列包含DDXXD保守序列,对其进行基因系统进化树分析表明,属于植物萜类合成酶基因家族Tps-a亚族。
因此,本发明还请求保护一种姜花倍半萜合成酶HcTPS12,其氨基酸序列如SEQ ID NO:3所示。
根据本发明提供的HcTPS12基因序列信息,本领域技术人员可以通过以下方法容易地获得与HcTPS12等同的基因:(1)通过数据库检索获得;(2)以HcTPS12基因片段为探针筛选姜花或其它植物的基因组文库或cDNA文库获得;(3)根据HcTPS12基因序列信息设计寡核苷酸引物,用PCR扩增的方法从姜花或其它植物的基因组、mRNA和cDNA中获取;(4)在HcTPS12基因序列的基础上用基因工程方法改造获得;(5)用化学合成的方法获得该基因。
本发明还提供一种用于扩增姜花倍半萜合成酶基因HcTPS12的引物对,所述引物对的核苷酸序列如SEQ ID NO:4~5所示。
同时,本发明还提供一种包含上述姜花倍半萜合成酶基因HcTPS12的重组载体。
一种包含上述重组载体的重组菌。
一种包含上述重组菌的细胞系。
本发明通过将姜花倍半萜合成酶基因HcTPS12的全长cDNA序列连接到原核表达载体上,经过体外诱导得到高纯度的体外重组蛋白。通过给予反应底物FPP对上述体外重组蛋白进行体外酶活实验。结果显示HcTPS12蛋白催化FPP生成倍半萜成分红没药烯,占反应总产物的100%。红没药烯为姜花精油组成成分之一。表明本发明的HcTPS12蛋白是具有单一催化功能的倍半萜合成酶,同时可利用姜花倍半萜合成酶基因HcTPS12通过微生物代谢工程来制备红没药烯,进而制备植物精油或药物。
因此,本发明所述姜花倍半萜合成酶基因HcTPS12和/或姜花倍半萜合成酶HcTPS12在制备红没药烯,或在制备香料、精油或药物中的应用也在本发明保护范围内。
具体地,所述应用为以法呢基焦磷酸(FPP)为底物,经姜花倍半萜合成酶HcTPS12催化产生红没药烯。
本发明提供的姜花倍半萜合成酶基因HcTPS12具有重要的应用价值。应用之一是将所述的HcTPS12基因序列连接到任何一种植物转化载体,用任何一种转化方法将HcTPS12基因导入姜花或其他植物细胞,可获得表达所述基因的转基因植株,从而应用于生产。本发明所述的基因构建到植物转化载体中时,可以对所述基因或其调控序列适当修饰,也可以在其转录起始密码子前用其它启动子取代所述基因原有的启动子,从而拓宽和增强植物产生倍半萜红没药烯和增强抗性的能力。
本发明提供的倍半萜合成酶基因HcTPS12的再一个应用是根据所述基因序列信息产生特异性的分子标记,包括但不限于SNP(单核苷酸多态)、SSR(简单序列重复多态)、RFLP(限制性内切酶长度多态)、CAP(切割扩增片段多态)。用这些标记可鉴定姜花或其它植物的倍半萜红没药烯合成酶基因,用于分子标记辅助选择育种,从而提高育种的选择效率。
与现有技术相比,本发明具有以下有益效果:
本发明提供了一种新的倍半萜合成酶基因HcTPS12,所述基因可以催化FPP形成倍半萜类化合物红没药烯,在提高植物萜类成分含量和抗性方面都发挥着重要作用;所述半萜合成酶基因HcTPS12可用于制备红没药烯及进一步制备精油、香精和药物。将本发明的基因片段构建在植物表达载体上,可以外源转化其他植物材料,从而获得含有倍半萜红没药烯合成酶基因的转基因材料,对培育药用植物提供了一种有效地方法。倍半萜合成酶基因的克隆是克服传统育种中不能在植物种间转移基因问题的前提。另外,本发明能够进一步提供或应用利用上述DNA片段获得的具有药用价值的转基因植株和相应的种子,以及用本发明的基因或基于该基因的重组体转化的植株或由这类植株获得的种子。可以用有性杂交的方式将本发明的基因转入其他的植株。
附图说明
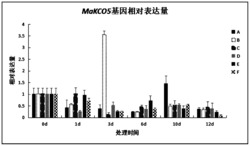
图1为本发明HcTPS12基因在姜花不同组织中表达特异性情况;Pe:花瓣;Ri:根茎;Le:叶片。
图2为本发明HcTPS12基因在姜花叶器官不同发育时期的表达情况。
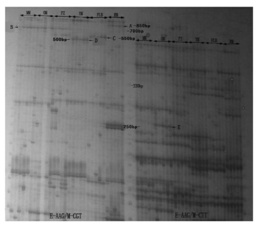
图3为本发明HcTPS12重组蛋白以FPP为底物体外酶催化反应。
具体实施方式
下面结合附图和具体实施例进一步详细说明本发明。除非特别说明,实施例中采用的试剂和方法均为本领域常规使用的试剂和方法。
实施例1 HcTPS12基因cDNA全长的获得
S1.姜花叶片RNA的提取:以超低温冰箱保存的姜花叶片作为提取RNA的材料。RNA提取所用的枪头、Eppendorf管用0.1%的DEPC于37℃浸泡过夜后121℃高温灭菌25min,玻璃器皿,研钵用铝箔包好后于180℃下干热处理3h,冷却后备用。采用Trizol法按照Trizol(TaKaRa)的说明书提取姜花叶片总RNA。用1%琼脂糖凝胶电泳检测RNA的完整性,利用微量分光光度计法测定其浓度和纯度。放于-80℃保存备用。
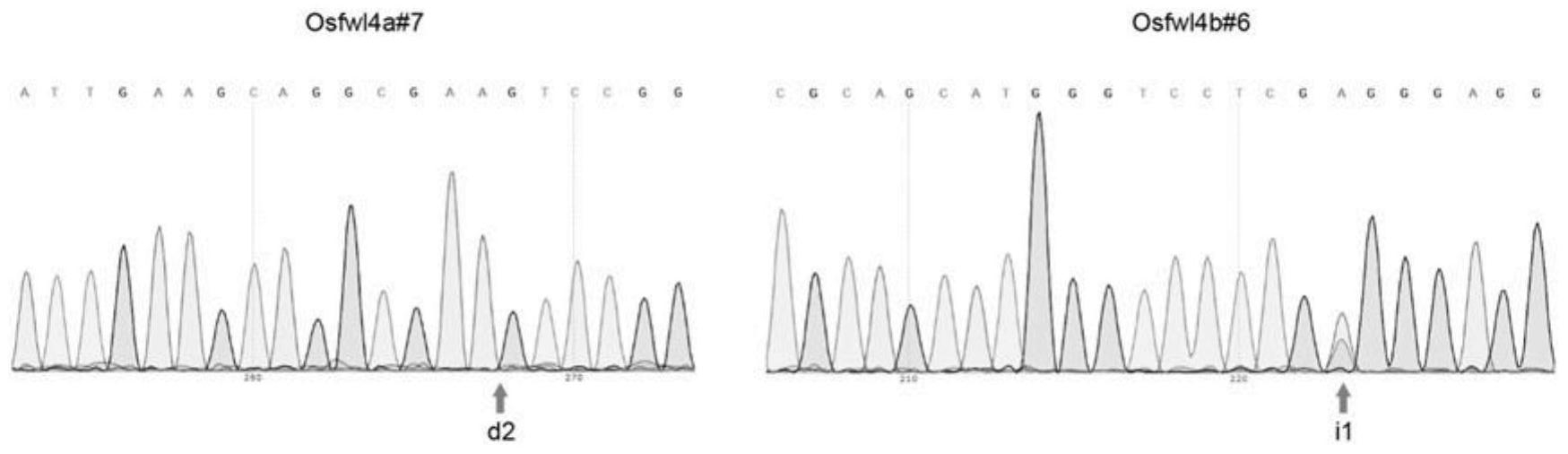
S2.姜花叶片总RNA作为模板,用生工M-MuLV First cDNA Synthesis Kit合成第一链cDNA。据姜花转录组数据库相关注释基因序列,设计引物,上游引物F1:5’-GTCATGGAGCTTGCTGGTACT-3’(如SEQ ID NO:4所示)。下游引物R1:5’-GGCTTCAACAATATTAACAAGACC-3’(如SEQ ID NO:5所示)。并交由上海生物工程公司合成。以上述合成的cDNA为模板采用TaKaRa PCR Amplification Kit进行PCR扩增反应,具体方法按照说明书进行。PCR程序为:94℃预变性4min;94℃变性30s、56℃复性30s、72℃延伸2min,35个循环;然后72℃延伸10min。在-20℃下保存备用。PCR反应结束后,用1.0%琼脂糖凝胶电泳初步检测PCR产物中是否含有目的片段条带。PCR扩增产物经1%琼脂糖凝胶电泳检测后,用手术刀在紫外灯下切出含有目的片段的胶块,用DNA凝胶回收试剂盒(Agarose Gel DNA Purification Kit,TaKaRa)进行回收,回收方法基本参照试剂盒说明书。之后要对回收产物进行1%琼脂糖电泳检测,看其回收效果及大致浓度,以便保证后续试验的进行。根据回收的目的片段大小及其有效浓度,取适量回收纯化的产物与克隆载体连接,载体选用TaKaRa pMD19-T载体,目的DNA与克隆载体的摩尔比控制在3:1左右,具体操作按说明书进行。在16℃下恒温连接3~6h,连接时间的长短视目的片段的长度而定。提前将感受态细胞DH5α(TaKaRa)从-80℃冰箱取出,并置于冰盒中待其自然融化,将10μL连接液全量加入感受态细胞的离心管中,冰浴30min后,42℃水浴热激50s,迅速冰上放置2~5min,然后加入37℃预温的SOC液体培养基890μL补足到1mL,混匀后37℃下180rpm振荡培养1h。在含100μg/mL氨苄青霉素的LB固体培养基平板表面上涂30μL的X-gal(20mg/mL)和30μL IPTG(20mg/mL),然后涂布适量转化液,待转化液完全被吸收后倒置于37℃恒温箱中过夜培养,约16h后观察结果,通过X-gal/IPTG蓝白斑筛选白色菌落,并初步鉴定重组质粒,平板置于4℃保存。通过蓝白斑初步筛选后,通常选6个菌斑摇菌后提取质粒,用于进一步鉴定。用灭菌的牙签从LB平板培养基上挑取白色单菌落接种于含有100μg/mL氨苄青霉素的LB液体培养基中,于控温震荡水浴摇床上37℃、240rpm振荡过夜培养,用质粒DNA小抽试剂盒(上海博亚生物有限公司)提取质粒,步骤按说明书上方法进行。对所提质粒进行1.0%琼脂糖凝胶电泳检测,比较质粒大小并将明显滞后的质粒进行双酶切(EcoRI/Hind III,TaKaRa)分析。在37℃下酶切1h后,对酶切产物进行1.0%琼脂糖凝胶电泳检测。随意选送酶切鉴定后含目的片段的重组质粒进行DNA序列测定。测序工作交由上海生工生物技术有限公司完成,采用美国ABI377序列分析仪。所得的序列和转录组原有序列信息进行比对,在NCBI进行比对和同源性分析,确定获得的基因序列为TPS家族的完整全长序列,同时根据cDNA序列推测出其蛋白序列。
结果所示HcTPS12基因的全长cDNA序列如SEQ ID NO:1所示,cDNA全长为1823bp;基因编码区(CDS)如SEQ ID NO:2所示,共1653bp;推测其氨基酸序列如SEQ ID NO:3所示。
实施例2 HcTPS12基因的表达分析
1、选取姜花不同组织部位及不同发育时期叶片提取RNA,其RNA提取均使用Trizol法(TaKaRa),荧光定量PCR采用SYBR green(TaRaKa)法,染料法的具体原理见说明书。利用Primer Premier 5.0软件设计实时荧光定量PCR引物,按荧光定量PCR引物设计原则,用Primer premier 5.0分别设计引物,通过荧光定量PCR检测其是否有错配或引物二聚体及其扩增效率,从中选择一对最佳引物,P1:5’-TGGAAGGCGTGGTTGTTGAT-3’(如SEQ ID NO:6所示)。P2:5’-AGACCGTGATTTCTGATTTGT-3’(如SEQ ID NO:7所示)。内参基因RPS根据Real-time PCR引物的设计原则,用Primer premier 5.0设计引物,RPS-P1:5’-TTAGTAGCATCGGCTGCAATAAG-3’(如SEQ ID NO:8所示),RPS-P2:5’-CTCAACCGTCTTCCCAAAAGAG-3’(如SEQ ID NO:9所示)。通过Real-time PCR进行检测,并制作标准曲线,以检测其扩增效率(E)是否在90~110%范围内进行筛选。以各样品的cDNA为模板,在ABI荧光定量PCR仪上进行荧光定量PCR反应。每个样品设3个重复,以ddH2O为阴性对照。反应体系为SYBR Premix ExTaq(TaKaRa)10.0μL,上游引物(10μM)0.4μL,下游引物(10μM)0.4μL,cDNA 2.0μL,ddH2O 7.2μL。反应程序为94℃,30s;94℃,15s;55℃,30s;72℃,30min;40个循环,94℃,15s;72℃,30s;0.4℃/s融解曲线分析。反应结束后确认扩增曲线和融解曲线,采用2-△△Ct法(Livak et al.,2001)进行数据分析,计算姜花HcTPS12在不同样品中的表达情况。
2、结果
基因表达分析结果见图1,2所示。从图1中可以看出:HcTPS12在叶组织中表达量较高,在根茎和叶等营养器官几乎不表达。图2表明:该基因的表达量受叶片发育的调控,在老叶中表达量更高,与姜花精油中倍半萜红没药烯含量的变化趋势一致,说明HcTPS12是参与调控姜花萜类物质合成的相关基因。
实施例3HcTPS12基因原核表达
S1.载体构建:根据所得到HcTPS12基因的编码区,用包含KpnI和EcoRI酶切位点的特异引物F:5’-GATCTGGGTACCATGGAGCTTGCTGGTACTCC-3’(如SEQ ID NO:10所示);R:5’-GAGCTCGAATTCAATAGGGATAGGATGAACAA-3’(如SEQ ID NO:11)所示。进行PCR扩增。PCR产物用Takara回收试剂盒回收,回收产物直接用KpnI和EcoRI限制性内切酶进行双酶切,1%琼脂糖凝胶回收目的片段。pET-30a原核表达载体用KpnI和EcoRI限制酶进行双酶切1%琼脂糖凝胶回收大片段。于16℃连接过夜,将连接产物转化大肠杆菌(E.coli)DH5α感受态细胞;提取质粒经酶切和测序鉴定后,获得重组原核表达载体。
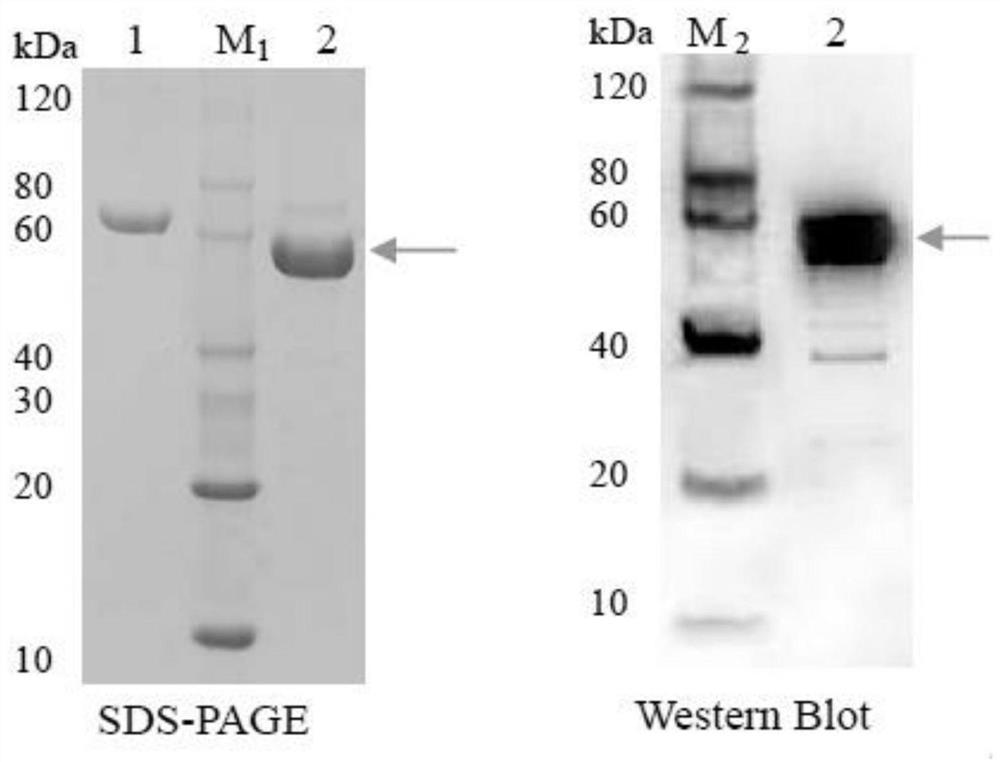
S2.重组蛋白表达:用经鉴定的重组质粒DNA转化BL21(DE3)感受态细胞,挑取单菌落接种于5mL LB(含25mg/L Kan,34mg/L Chl)培养基中,37℃震荡培养过夜。转接100μL种子液于新鲜的100mL(含25mg/L Kan,34mg/L Chl)的LB培养基中,37℃ 180rpm培养4~6h,测OD值至0.4~0.6后用一定量的IPTG(0.1~0.2mM)在14~18℃诱导14~16h。同时取另一对照组其中未加IPTG诱导。离心收集菌体,用5mL裂解buffer(50mM磷酸buffer pH 8.0)悬浮细胞,将菌体冷却,置于冰上超声波破碎细胞。12000rpm,4℃离心10min,将上清移至新的离心管中,用双蒸水清洗沉淀一次,再用5mL裂解buffer悬浮沉淀。分别取上清和沉淀各50μL,保存与-20℃,进行SDS-PAGE电泳分析。配制12.5%SDS聚丙烯酰胺凝胶,按照顺序上样。分别用60V和120V的电压对浓缩胶和分离胶的进行电泳。电泳完毕,考马斯亮蓝染色30min,再用脱色液脱色1~8h,观察并记录试验结果。
S3.重组蛋白的纯化:挑取单菌落接种于5mL的液体LB培养基中,37℃,180rpm,培养过夜,后全部转接至500mL的新鲜液体LB培养集中,37℃,180rpm,培养4h,用IPTG(0.1~0.2mM),18℃条件下诱导16h后,收集菌体。取200μL菌体于离心管中,4℃保存,进行SDS-PAGE电泳分析。用5mL的裂解buffer悬浮细胞并转移至离心管内,置于冰上使菌体始终保持冷却,超声波破碎细胞。10,000g,4℃离心20~30min,收集上清液。取20μL的上清于-20℃保存,进行SDS-PAGE分析。向5mL细胞裂解液中加入1~2mL的Ni-NTA resin(镍--次氮基三乙酸树脂填料)混匀,并低速在4℃摇床上结合60min。将结合完全的细胞裂解液和树脂混合物装入层析柱中,移走底部盖帽收集流出液部分(flow-through fraction,F),取20μL的流出液,于-20℃保存,进行SDS-PAGE分析。用4mL的洗涤buffer洗脱层析柱两次,收集每部分的洗脱液(wash fraction,W),各取20μL保存于-20℃冰箱中作SDS-PAGE分析。用0.5mL的洗脱buffer洗柱四次,分别用不同的收集管依次收集每部分的洗脱液(elution fraction,E),分别标记为E1,E2,E3,E4,各取20μL进行SDS-PAGE分析。根据SDS-PAGE的检测结果,集中含有目标蛋白洗脱部分于一个离心管中,用移液器转移至透析袋中,4℃透析过夜。收集透析后的溶液,加入甘油使蛋白溶液中甘油总含量为20%,并且以200μL/管的量进行分装,取微量进行SDS-PAGE蛋白浓度检测,其余的放于-80℃超低温冰箱进行保存备用。
S4.酶催化功能鉴定:将30mM HEPES,100μL pH 7.5,5mM DTT,20mM MgCl2各100μL,以及蛋白提取液100μL,2μL GPP或者FPP,加ddH2O 588μL,使终体积为1mL,加入样品瓶中密封,30℃水浴1h,将75μm聚二甲基氧烷(PMDS)萃取纤维头插入玻璃瓶中,顶空固相微萃取1h,反应结束后将萃取纤维头放至气相色谱-质谱连用仪中进行分析,气相色谱条件为:色谱柱为HP-1NNOWAX柱(30m×0.25mm);载气为高纯氦气,分流比20:1,柱前压50Pa,流量1mL/min;取样时间2min;程序升温:柱起始温度45℃,保持2min,以5℃/min的速度升至80℃保持1min,然后以10℃/min的速度升至250℃保持5min。质谱条件为:GC-MS的接口温度220℃,电子轰击源EI、350V;离子源温度170℃;电子能量70eV;扫描质量范围35~335amu,采集到的质谱图用WILLEY/MAINLIB库进行分析。
HcTPS12基因原核表达结果见图3。图3所示:以FPP为底物时,pET-30a-HcTPS12体外酶催化反应产物经质谱鉴定为倍半萜类物质红没药烯。说明HcTPS12基因编码生成的酶是一种可以催化FPP的生成半萜类物质红没药烯的单功能酶基因。
序列表
<120> 一种姜花倍半萜合成酶基因HcTPS12及其应用
<141> 2019-02-28
<160> 11
<170> SIPOSequenceListing 1.0
<210> 1
<211> 1823
<212> DNA
<213> 姜花(Hedychium coronarium Koen)
<400> 1
caagtcatgg agcttgctgg tactccatca gtcgaggttt tggaaggcgt ggttgttgat 60
cgtcagttgg caggcttcga tcccagctct tggggtgaat actttattac aaatcagaaa 120
tcacggtctg aggcatggat gaacgaaaga gctgaagagc tcaagaatga agtaaggagc 180
atgttccaaa acgtgactgg cgtcctacaa accatgaatc taattgatac aattcaactc 240
ctcggacttg attaccattt catggaggaa atagacagag ctttagatca tctcaaggat 300
gttgacatga gcaaaaacgg gctctatgag gttgctcttc attttcgact gcttagacaa 360
aaaggagtca acatttcttc agatgtattt aaaaaataca tggataaaga gggaaaattt 420
atagaagaac taaaagatga tgctaagggg ctcctgagct tatataatgc ggcttacctc 480
ggaaccaaaa aagagactat actcgacgaa gccatttctt ttactagaga taaccttaca 540
tctttgttaa aagatctaaa tcctacattt gcaaagttag tgtctctcac tctaaagaca 600
cctattccac gaagcatgaa acgacttttc acaagatgct acatctctat ttaccaagat 660
gaaccgactc gaaatgaaac aatatttgag cttgcaaaat tggacttcaa catactacaa 720
tgtctccacc aggaggagct caagaaacta agcatgtggt ggaagcagtt gaatttagac 780
atcatgcatc taaattttgt tcgagatcga gtggtggaat gtttctgttg gtcgatggag 840
atacgccatg aacccagttg ttctcgtgct cgactgatag cgtctaagct acttatgttc 900
attactgtct tggatgactt ctatgatagc tacagcacat tagaagagag tcgactactt 960
acagatgcaa tcgaaaggtg gagtcctgat gcagtagatc aactaccaga atacctgagg 1020
gagttctttc tcaaaatgtt gaacattttt caagaatttg aagatgaact tgcaccggaa 1080
gagaagtttc gaatattgta cttcaaggaa gaatggaaaa ctcaagctca aagttacttc 1140
aaggaatgcc aatggaggga tgacaattat gtgcccaagt tagaagagca catgcgtgtt 1200
tcaatcataa gtgtgggatt tgtcttgttt tattgcggat ttttgagtgg catggaggag 1260
acagtggcca caaaggatgc atttgaatgg ttcgcaagct tccccaagat catagaagct 1320
tgtgcaacaa ttcttcgtat cactaatgac ataacttcaa aggagcgaga acaaaagagg 1380
gcacatgttg cctcgacggt agattgctat atgaaggaat atggaacatc aaaagatgtt 1440
gcatgcgaga agctcctagg ctttgttgaa gatgcatgga agactatcaa cgaggagctc 1500
cttactgcaa ctggattgtc gagggaagta attgaactat cactccactg tgcgcaaact 1560
acagaatttg tatacaagga tgtcgacgca ttcacagaac ctaatacctc gatgaaggaa 1620
agcatctttt tcctacttgt tcatcctatc cctatttgat gacagtagtg ctacaatcat 1680
gtactatttg gtatctcata tggttgtgtg cttaagttat tattatagaa ataaaagggg 1740
ggagaaagag agaaataact tggtcttgtt aatattgttg aagccaaata agcttatatg 1800
tacaagttgt ttaccagttg tac 1823
<210> 2
<211> 1653
<212> DNA
<213> 姜花(Hedychium coronarium Koen)
<400> 2
atggagcttg ctggtactcc atcagtcgag gttttggaag gcgtggttgt tgatcgtcag 60
ttggcaggct tcgatcccag ctcttggggt gaatacttta ttacaaatca gaaatcacgg 120
tctgaggcat ggatgaacga aagagctgaa gagctcaaga atgaagtaag gagcatgttc 180
caaaacgtga ctggcgtcct acaaaccatg aatctaattg atacaattca actcctcgga 240
cttgattacc atttcatgga ggaaatagac agagctttag atcatctcaa ggatgttgac 300
atgagcaaaa acgggctcta tgaggttgct cttcattttc gactgcttag acaaaaagga 360
gtcaacattt cttcagatgt atttaaaaaa tacatggata aagagggaaa atttatagaa 420
gaactaaaag atgatgctaa ggggctcctg agcttatata atgcggctta cctcggaacc 480
aaaaaagaga ctatactcga cgaagccatt tcttttacta gagataacct tacatctttg 540
ttaaaagatc taaatcctac atttgcaaag ttagtgtctc tcactctaaa gacacctatt 600
ccacgaagca tgaaacgact tttcacaaga tgctacatct ctatttacca agatgaaccg 660
actcgaaatg aaacaatatt tgagcttgca aaattggact tcaacatact acaatgtctc 720
caccaggagg agctcaagaa actaagcatg tggtggaagc agttgaattt agacatcatg 780
catctaaatt ttgttcgaga tcgagtggtg gaatgtttct gttggtcgat ggagatacgc 840
catgaaccca gttgttctcg tgctcgactg atagcgtcta agctacttat gttcattact 900
gtcttggatg acttctatga tagctacagc acattagaag agagtcgact acttacagat 960
gcaatcgaaa ggtggagtcc tgatgcagta gatcaactac cagaatacct gagggagttc 1020
tttctcaaaa tgttgaacat ttttcaagaa tttgaagatg aacttgcacc ggaagagaag 1080
tttcgaatat tgtacttcaa ggaagaatgg aaaactcaag ctcaaagtta cttcaaggaa 1140
tgccaatgga gggatgacaa ttatgtgccc aagttagaag agcacatgcg tgtttcaatc 1200
ataagtgtgg gatttgtctt gttttattgc ggatttttga gtggcatgga ggagacagtg 1260
gccacaaagg atgcatttga atggttcgca agcttcccca agatcataga agcttgtgca 1320
acaattcttc gtatcactaa tgacataact tcaaaggagc gagaacaaaa gagggcacat 1380
gttgcctcga cggtagattg ctatatgaag gaatatggaa catcaaaaga tgttgcatgc 1440
gagaagctcc taggctttgt tgaagatgca tggaagacta tcaacgagga gctccttact 1500
gcaactggat tgtcgaggga agtaattgaa ctatcactcc actgtgcgca aactacagaa 1560
tttgtataca aggatgtcga cgcattcaca gaacctaata cctcgatgaa ggaaagcatc 1620
tttttcctac ttgttcatcc tatccctatt tga 1653
<210> 3
<211> 550
<212> PRT
<213> 姜花(Hedychium coronarium Koen)
<400> 3
Met Glu Leu Ala Gly Thr Pro Ser Val Glu Val Leu Glu Gly Val Val
1 5 10 15
Val Asp Arg Gln Leu Ala Gly Phe Asp Pro Ser Ser Trp Gly Glu Tyr
20 25 30
Phe Ile Thr Asn Gln Lys Ser Arg Ser Glu Ala Trp Met Asn Glu Arg
35 40 45
Ala Glu Glu Leu Lys Asn Glu Val Arg Ser Met Phe Gln Asn Val Thr
50 55 60
Gly Val Leu Gln Thr Met Asn Leu Ile Asp Thr Ile Gln Leu Leu Gly
65 70 75 80
Leu Asp Tyr His Phe Met Glu Glu Ile Asp Arg Ala Leu Asp His Leu
85 90 95
Lys Asp Val Asp Met Ser Lys Asn Gly Leu Tyr Glu Val Ala Leu His
100 105 110
Phe Arg Leu Leu Arg Gln Lys Gly Val Asn Ile Ser Ser Asp Val Phe
115 120 125
Lys Lys Tyr Met Asp Lys Glu Gly Lys Phe Ile Glu Glu Leu Lys Asp
130 135 140
Asp Ala Lys Gly Leu Leu Ser Leu Tyr Asn Ala Ala Tyr Leu Gly Thr
145 150 155 160
Lys Lys Glu Thr Ile Leu Asp Glu Ala Ile Ser Phe Thr Arg Asp Asn
165 170 175
Leu Thr Ser Leu Leu Lys Asp Leu Asn Pro Thr Phe Ala Lys Leu Val
180 185 190
Ser Leu Thr Leu Lys Thr Pro Ile Pro Arg Ser Met Lys Arg Leu Phe
195 200 205
Thr Arg Cys Tyr Ile Ser Ile Tyr Gln Asp Glu Pro Thr Arg Asn Glu
210 215 220
Thr Ile Phe Glu Leu Ala Lys Leu Asp Phe Asn Ile Leu Gln Cys Leu
225 230 235 240
His Gln Glu Glu Leu Lys Lys Leu Ser Met Trp Trp Lys Gln Leu Asn
245 250 255
Leu Asp Ile Met His Leu Asn Phe Val Arg Asp Arg Val Val Glu Cys
260 265 270
Phe Cys Trp Ser Met Glu Ile Arg His Glu Pro Ser Cys Ser Arg Ala
275 280 285
Arg Leu Ile Ala Ser Lys Leu Leu Met Phe Ile Thr Val Leu Asp Asp
290 295 300
Phe Tyr Asp Ser Tyr Ser Thr Leu Glu Glu Ser Arg Leu Leu Thr Asp
305 310 315 320
Ala Ile Glu Arg Trp Ser Pro Asp Ala Val Asp Gln Leu Pro Glu Tyr
325 330 335
Leu Arg Glu Phe Phe Leu Lys Met Leu Asn Ile Phe Gln Glu Phe Glu
340 345 350
Asp Glu Leu Ala Pro Glu Glu Lys Phe Arg Ile Leu Tyr Phe Lys Glu
355 360 365
Glu Trp Lys Thr Gln Ala Gln Ser Tyr Phe Lys Glu Cys Gln Trp Arg
370 375 380
Asp Asp Asn Tyr Val Pro Lys Leu Glu Glu His Met Arg Val Ser Ile
385 390 395 400
Ile Ser Val Gly Phe Val Leu Phe Tyr Cys Gly Phe Leu Ser Gly Met
405 410 415
Glu Glu Thr Val Ala Thr Lys Asp Ala Phe Glu Trp Phe Ala Ser Phe
420 425 430
Pro Lys Ile Ile Glu Ala Cys Ala Thr Ile Leu Arg Ile Thr Asn Asp
435 440 445
Ile Thr Ser Lys Glu Arg Glu Gln Lys Arg Ala His Val Ala Ser Thr
450 455 460
Val Asp Cys Tyr Met Lys Glu Tyr Gly Thr Ser Lys Asp Val Ala Cys
465 470 475 480
Glu Lys Leu Leu Gly Phe Val Glu Asp Ala Trp Lys Thr Ile Asn Glu
485 490 495
Glu Leu Leu Thr Ala Thr Gly Leu Ser Arg Glu Val Ile Glu Leu Ser
500 505 510
Leu His Cys Ala Gln Thr Thr Glu Phe Val Tyr Lys Asp Val Asp Ala
515 520 525
Phe Thr Glu Pro Asn Thr Ser Met Lys Glu Ser Ile Phe Phe Leu Leu
530 535 540
Val His Pro Ile Pro Ile
545 550
<210> 4
<211> 21
<212> DNA
<213> 姜花(Hedychium coronarium Koen)
<400> 4
gtcatggagc ttgctggtac t 21
<210> 5
<211> 24
<212> DNA
<213> 姜花(Hedychium coronarium Koen)
<400> 5
ggcttcaaca atattaacaa gacc 24
<210> 6
<211> 20
<212> DNA
<213> 姜花(Hedychium coronarium Koen)
<400> 6
tggaaggcgt ggttgttgat 20
<210> 7
<211> 21
<212> DNA
<213> 姜花(Hedychium coronarium Koen)
<400> 7
agaccgtgat ttctgatttg t 21
<210> 8
<211> 23
<212> DNA
<213> 姜花(Hedychium coronarium Koen)
<400> 8
ttagtagcat cggctgcaat aag 23
<210> 9
<211> 22
<212> DNA
<213> 姜花(Hedychium coronarium Koen)
<400> 9
ctcaaccgtc ttcccaaaag ag 22
<210> 10
<211> 32
<212> DNA
<213> 姜花(Hedychium coronarium Koen)
<400> 10
gatctgggta ccatggagct tgctggtact cc 32
<210> 11
<211> 32
<212> DNA
<213> 姜花(Hedychium coronarium Koen)
<400> 11
gagctcgaat tcaataggga taggatgaac aa 32
一种姜花倍半萜合成酶基因HcTPS12及其应用专利购买费用说明
![]()
Q:办理专利转让的流程及所需资料
A:专利权人变更需要办理著录项目变更手续,有代理机构的,变更手续应当由代理机构办理。
1:专利变更应当使用专利局统一制作的“著录项目变更申报书”提出。
2:按规定缴纳著录项目变更手续费。
3:同时提交相关证明文件原件。
4:专利权转移的,变更后的专利权人委托新专利代理机构的,应当提交变更后的全体专利申请人签字或者盖章的委托书。
Q:专利著录项目变更费用如何缴交
A:(1)直接到国家知识产权局受理大厅收费窗口缴纳,(2)通过代办处缴纳,(3)通过邮局或者银行汇款,更多缴纳方式
Q:专利转让变更,多久能出结果
A:著录项目变更请求书递交后,一般1-2个月左右就会收到通知,国家知识产权局会下达《转让手续合格通知书》。









动态评分
0.0