

IPC分类号 : C07K16/28,C07K14/705,C07K17/00,C12N5/10,C12N1/15,C12N1/19,C12N1/21,C12N15/12,C12Q1/68,A61K39/395,A61P35/00,G01N33/68,G01N33/53







专利摘要
本发明提供了分离的多肽,其氨基酸序列与SEQ ID NO:20具有至少70%同一性,其中该多肽具有ER-α36活性。本发明还提供了鉴别能结合这样的多肽的试剂的方法,检测这样的多肽的方法,和改变这样的多肽的活性的方法。还提供了能特异性地结合SEQ ID NO:1所述的氨基酸序列,或其免疫原性片段的抗体,和制备和使用这样的抗体的方法。
说明书
本申请是申请号为200580007467.8(PCT/US2005/007857),申请日为2005年3月10日的中国发明专利申请“雌激素受体和使用方法”的分案申请。
继续申请数据
本申请要求享有2004年3月10日提交的美国临时申请系列号60/552,067和2005年1月13日提交的60/643,469的利益,其每篇都在这里引作参考。
政府资助
本文所述的发明是在健康和人类服务部(Department of Health and Human Services)资助号CA84328的支持下完成的。美国政府享有本发明的某些权利。
发明背景
雌激素是在卵巢、睾丸和可能的肾上腺皮质中形成的类固醇化合物的总称。雌激素和具有雌激素活性的化合物的实例包括二乙基己烯雌酚、己烯雌酚二磷酸酯、己雌酚、聚磷酸雌二醇、溴苯雌酚、氯烯雌醚、己二烯雌酚、二乙基己烯雌酚、丙甲雌酚、双醋羟雌酮、6,9-去氢马烯雌酮、马烯雌酮、雌二醇、雌三醇、雌酮、乙炔雌二醇、美雌醇、mexestrol、雌三醇环戊醚和炔雌醚。雌激素调节生殖组织和乳房、心血管、骨、肝和脑组织中的多种生理过程。雌激素也用在口服避孕药中。雌激素的其它用途包括,缓解绝经的不适,抑制泌乳,和治疗骨质疏松症、先兆流产和各种功能性卵巢病症。抗雌激素药被用于治疗转移性乳腺癌和晚期前列腺癌。
雌激素的作用由雌激素受体介导。在1986年克隆了首个雌激素受体(ER)(Green等,Nature,320:134(1986)和Greene等,Science,231:1150(1986))。在1995年之前,认为仅有一个负责天然的和合成的雌激素和抗雌激素药的所有生理和药理作用的雌激素受体。然而,在1995年,克隆了第二个雌激素受体(Kuiper等,PNAS,93:5925(1996))。发现的首个雌激素受体现在称作雌激素受体-α(ER-α),而第二个雌激素受体称作雌激素受体-β(ER-β)。
ER-α和ER-β共有共同的结构体系(Zhang等,FEBS Letters,546:17(2003)和Kong等,Biochem.Soc.Trans.,31:56(2003))。二者都由3个独立的但是能相互作用的功能结构域组成:N-末端A/B结构域、C或DNA-结合结构域和D/E/F或配体-结合结构域(图1)。ER-α的N-末端结构域编码独立于配体的激活功能(AF-1),即参加与辅激活物的相互作用和靶基因的转录活化的区域。DNA-结合结构域或C结构域含有2个锌指结构,其在受体二聚化和与特定的DNA序列的结合中起重要作用。C-末端D/E/F结构域是一种配体-结合结构域,其能介导配体结合、受体二聚化、核转位和配体-依赖性的反式激活功能(AF-2)。AF-1和AF-2对转录控制的相对贡献以细胞特异性的和DNA 启动子特异性的方式变化(Berry等,EMBO J.,9:2811(1990)和Tzukerman等,Mol.Endocrin.,8:21(1994))。
已经表明,缺少ER-α基因的全长基因产物的前173个氨基酸的46-kDa ER-α同种型(A/B或AF-1结构域),源自跳过外显子1的ER-α基因的可变剪接(Flouriot等,EMBO J.,19:4688(2000))。该可变剪接事件会产生mRNA,其在有利于翻译起始的Kozak序列内具有与原始可读框的剩余部分符合读框的AUG。因此,ER-α的该新同种型称作ER-α46,而原始同种型称作ER-α66(Flouriot等,EMBOJ.,19:4688(2000))。ER-α46形成同二聚体,并结合雌激素反应元件(ERE),且它也可以与ER-α66形成异二聚体(Flouriot等,EMBO J.,19:4688(2000))。与ER-α66同二聚体相比,ER-α46同二聚体表现出更高的对ERE的亲和力。而且,ER-α46/66异二聚体比ER-α66同二聚体优先形成,且ER-α46竞争地起作用,以抑制由结合了配体的-ER-α66的AF-1结构域介导的反式激活,但是不影响AF-2-依赖性的反式激活(Floutiot等,EMBO J.,19:4688(2000))。因此,认为ER-α46是天然产生的ER-α同种型,其调节由ER-α66的AF-1结构域介导的雌激素信号传导。
ER-α能在大约15-30%内腔(luminal)上皮细胞中表达,但是根本不在正常人乳房中的任何其它细胞类型中表达。双标记免疫荧光技术揭示,在正常的人和啮齿动物乳腺中,ER-α-表达细胞能与用增殖标记物标记的细胞分开(Clarke等,Cancer Res.,57:4987(1997))。在导管增生的最早期,ER-α表达会增加,甚至随着异型的增加而增加更多,从而在低和中等核级的不典型导管增生和原位导管癌中的大部分细胞含有ER-α(Khan等,Cancer Res.,54:993(1994)和Lawson等,Lancet,351:1787(1994))。随着ER-α表达的增加,受体表达和细胞增殖之间的反相关变得调节异常(Shoker等,Amer.Jour.Path.,155:1811(1999))。大约70%的侵入性乳腺癌表达ER-α,且这些肿瘤中的大部分含有ER-α-阳性的增殖细胞(Clarke等,Cancer Res.,57:4987(1997))。
雌激素受体是能控制许多生理过程的配体-活化的转录因子核受体超家族的成员。该控制经常通过调节基因转录来发生(Katzenellenbogen和Katzenellenbogen,Breast Cancer Res.,2:335(2000);Hull等,J.Biol.Chem.,276:36869(2001);McDonnell和Norris,Science,296:1642(2002))。雌激素受体使用多种机理,以激活或抑制它的靶基因的转录。这些机理包括:(a)在雌激素反应元件处,配体-占据的受体与DNA的直接相互作用,随后是转录辅调节物(coregulator)或介质复合物的募集,(b)配体-占据的ER与其它转录因子相互作用,所述其它转录因子例如AP-1(Kushner等,J.Steroid Biochem.Mol.Biol.,74:311(2000))、Sp1(Safe,Vitam.Horm.,62:231(2001))或NF-κB(McKay和Cidlowski,Endocr.Rev.,20:435(1999)),或(c)通过隔离一般的/普通的转录组分,间接调节基因转录(Harnish等,Endocrinology,141:3403(2000)和Speir等,Circ.Res.,87:1006(2000))。另外,雌激素受体通过这些不同的机理调节转录的能力,似乎是细胞类型特异性的,可能是由于在每种细胞类型中可得到的转录辅调节因子的补体的差异(Cerillo等,J.Steroid Biochem.Mol.Biol.,67:79(1998);Evans等,Circ.Res.,89:823(2001);Maret等,Endocrinology,140:2876(1999))。同样,转录调节依赖于配体的性质,其中各种天然的和合成的选择性的雌激素受体调节剂,通过这些不同机理中的每一种,起雌激素受体激动剂或拮抗剂的作用(Shang和Brown,Science,295:2465(2002);Katzenellenbogen和Katzenellenbogen,Science,295:2380(2002);Margeat等,J.Mol.Biol.,326:77(2003);Dang等,J.Biol.Chem.,278:962(2003))。
存在由雌激素介导的另一个信号传导途径,也称作“非经典的”、“非基因组的”或“膜信号传导”途径,其包含胞质蛋白、生长因子和其它膜-启动的信号传导途径(Segars等,Trends Endocrin.Met.,13:349(2002))。已经表明,几种细胞内的信号传导途径能与快速的雌激素-启动的作用通讯:腺苷酸环化酶途径(Aronica等,PNAS,91:8517(1994))、磷脂酶C途径(Le Mellay等,J.Cell.Biochem.,75:138(1999))、G-蛋白-偶联的受体-活化的途径(Razandi等,Mol.Endocrin.,13:307(1999))和促分裂原活化蛋白激酶(MAPK)途径(Watters等,Endocrinology,138:4030(1997))。但是,迄今为止描述的所有膜形式,都与ER-α有关,但与ER-β无关(Segars等,Trends Endocrin.Met.,13:349(2002))。
病理上,雌激素信号传导已经与乳腺癌和子宫内膜癌的高风险相关联(Summer和Fuqua,Semin.Cancer Biol.,11:339(2001);Turner等,Endocr.Rev.,15:275(1994);Farhat等,FASEB J.,10:615(1996);Beato等,Cell,83:851(1995);Dobrzycka等,Endo.Rel.Cancer,10:517(2003))。结果,已经发现雌激素受体是大多数这样的癌症的开始和发展所必需的。雌激素受体-阳性的乳腺癌的当前内分泌疗法主要针对雌激素水平、雌激素受体水平或雌激素和雌激素受体的活性设计。在早期乳腺癌的管理中,部分的抗雌激素药他莫昔芬的使用,已经清楚地证实了无疾病和总存活的增加。另外,近期的研究证实,他莫昔芬可以用作激素-依赖性的乳腺癌的化学预防剂。使用他莫昔芬的长期疗法的主要问题是它的亲子宫作用,这会导致子宫内膜癌的高风险,和对他莫昔芬的获得性临床抗性。这已经导致了对更好的选择性雌激素受体调节剂(SERM)的迫切寻求,所述调节剂在各种雌激素反应靶组织中表现出最佳的激动或拮抗活性。
因此,需要可以用于筛选能调节雌激素信号传导的其它方法和材料,以及可以用于调节雌激素信号传导的方法和材料。
发明概述
本发明提供了分离的抗体,其能特异性地结合SEQ ID NO:1所述的氨基酸序列,或其免疫原性片段,优选地,SEQ ID NO:1的氨基酸13-27所述的氨基酸序列。抗体可以是单克隆抗体或多克隆抗体。任选地,抗体是人源化的抗体。抗体可以共价地附着化合物,例如,化疗剂或检测标记例如荧光标记。抗体可以存在于组合物中,且组合物可以包含药学上可接受的载体。还提供了试剂盒,其包含本发明的抗体。
本发明也提供了制备抗体的方法。抗体可以是多克隆的或单克隆的。该方法包括,给动物施用具有SEQ ID NO:1所述的氨基酸序列的多肽,或其免疫原性片段,优选地,SEQ ID NO:1的氨基酸13-27所述的氨基酸序列。该方法还包括从动物分离抗体,其中该分离的抗体能特异性地结合氨基酸序列。多肽或其免疫原性亚基可以共价地附着载体多肽。分离可以包括,从动物得到产生该抗体的细胞,和使用该细胞制备能产生单克隆抗体的杂交瘤。本发明还包括通过该方法生产的多克隆抗体和通过该方法生产的单克隆抗体。
本发明也涉及包含外源编码区的细胞,其中该编码区编码包含SEQ ID NO:20的多肽。编码区可以编码具有与SEQ ID NO:20具有至少90%同一性的氨基酸序列的多肽,其中该多肽具有ER-α36活性。编码区可以可操作地连接到组成型启动子。细胞可以是真核细胞或原核细胞。本发明还提供了表达这样的多肽的细胞。
本发明还提供了鉴别结合多肽的试剂的方法。该方法包括,组合包含SEQ ID NO:1所述的氨基酸序列的多肽和试剂,检测试剂和多肽之间的复合物形成,检测多肽活性的改变,或其组合。通过直接检测试剂与多肽的结合,使用竞争结合测定检测试剂与多肽的结合,或其组合,可以检测试剂与多肽的结合。任选地,该方法也包括,确定试剂是否结合包含SEQ ID NO:18的多肽。
本发明还提供了检测多肽的方法。一方面,该方法包括提供细胞,分析细胞中具有ER-α36活性和36kDa分子量的多肽,所述分子量如按照在十二烷基硫酸钠(SDS)-聚丙烯酰胺凝胶上电泳测得的,和确定细胞是否表达该多肽。细胞可以是离体的(ex vivo)或体内的。细胞可以是,例如,肿瘤细胞,例如乳腺肿瘤细胞。分析可以包括,使细胞接触能特异性地结合SEQ ID NO:1所述的氨基酸序列,或其免疫原性片段的抗体。分析可以包括,扩增mRNA多核苷酸,以形成扩增的多核苷酸。扩增包括,使从细胞得到的多核苷酸接触引物对,后者能扩增mRNA多核苷酸,其包括SEQ ID NO:22或SEQ ID NO:25,或其组合,其中扩增的多核苷酸的存在表明该细胞表达多肽。引物对的一个引物可以选自SEQ ID NO:22的核苷酸,与SEQ ID NO:25的核苷酸互补的核苷酸,或其组合。
本发明也提供了抑制细胞的ER-α36活性的方法。该方法包括,使表达具有SEQ ID NO:1所述的氨基酸序列的多肽的细胞接触能抑制ER-α36活性的化合物。这样的化合物可以是能特异性地结合具有SEQ ID NO:1的氨基酸13-27所述的氨基酸序列的多肽的抗体。细胞可以是体内的或离体的,且任选地可以是ER-α66阴性的,ER-α46阴性的,或其组合。在有些方面,化合物不是抗雌激素药。
本发明还提供了分离的多肽,其包含SEQ ID NO:1的氨基酸13-27所述的氨基酸序列,优选地SEQ ID NO:1所述的氨基酸序列,更优选地,SEQ ID NO:20所述的氨基酸序列。在另一个方面,分离的多肽与SEQ ID NO:20具有至少70%同一性,其中该多肽具有ER-α36活性。本发明也包括SEQ ID NO:1的免疫原性片段。
当这些术语出现在说明书和权利要求书中时,术语“包含”和其变体不具有限制含义。除非另有限定,可互换地使用“一个(a)”、“一个(an)”、“该(the)”和“至少一个”,且指一个或多于一个。
附图简述
图1解释了人雌激素受体-α(ER-α)同种型的结构域结构代表。显示了结构域(用A-F标记的)、氨基酸序列编号、AF-1和AF-2、DNA结合结构域、配体-结合结构域和二聚化结构域。也指示了磷酸化位点和每个结构域的功能。
图2是证实ER-α的膜和基因组信号传导途径之间的可能的通讯的示意图。Cav-1代表小窝蛋白-1,ER-α,雌激素受体-α;RTK,受体酪氨酸激酶;Ras,Ras癌基因;Mek,MAP/ERK激酶;MAPK,促分裂原活化蛋白激酶;PI3K,磷酸肌醇-三磷酸激酶;AKT,蛋白激酶13;PDK1,磷酸肌醇-依赖性的蛋白激酶;RSK,p90核糖体S6激酶。
图3是显示在有雌二醇(E2)存在下,pRET-感染的MCF10A细胞在软琼脂中生长成大菌落的照片。ST1克隆在含有E2的软琼脂中表现出快速生长。分别包括MCF7和MCF10A细胞作为阳性和阴性对照。
图4是显示pRET-感染的MCF10A细胞中的小窝蛋白-1(Cav-1)表达的下调的蛋白印迹。通过使用兔抗-Cav-1抗体(N20)的蛋白印迹,分析来自不同细胞系的等量的全细胞提取物。用箭标指示Cav-1的位置,且在每个泳道上面指示在每个泳道中分析的细胞提取物。
图5是显示pRET-感染的MCF10A细胞中的ER-α表达的下调的蛋白印迹。通过使用针对ER-α(H222)和ER-β的抗体的蛋白印迹,分析来自不同细胞系的等量的全细胞提取物。用箭标指示ER-α和ER-β的位置,且在每个泳道上面指示在每个泳道中分析的细胞提取物。
图6是显示pRET-感染的MCF10A细胞中的ERK1/2磷酸化的活化的蛋白印迹。通过使用针对ERK1/2和磷酸化的ERK1/2的抗体的蛋白印迹,分析来自细胞系的等量的全细胞提取物。
图7是显示3种ER-α蛋白在Cav-1单倍体不足(haploinsufficient)细胞ST1和ST3和MCF7乳腺癌细胞中的存在的蛋白印迹。通过使用针对ER-α的H222抗体的蛋白印迹,分析来自细胞系的等量的全细胞提取物。用箭标指示ER-α66、ER-α46和ER-α36的位置,且在每个泳道上面指示在每个泳道中分析的细胞提取物。
图8描绘了人ER-α基因的基因组组织。用箭标指示多个启动子的位置。用AUG和UGA指示翻译起始和终止位置。用编号的框显示外显子。也用框中的外显子1′显示内含子1。下图显示了ER-α同种型的mRNA结构。用AAA指示聚腺苷酸化位点。
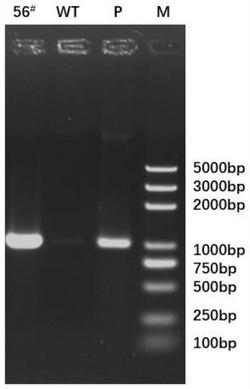
图9是琼脂糖凝胶的图片,它显示了通过PCR分离能编ER-α36的可读框的cDNA。用箭标指示cDNA在凝胶中的位置。
图10显示了ER-α36可读框的预测的氨基酸序列。用氨基酸序列(SEQ ID NO:20)左侧的编号,指示氨基酸位置。ER-α36独有的最后的27个氨基酸标有下划线(SEQ ID NO:1)。
图11显示了ER-α66、ER-α46和ER-α36的蛋白印迹分析。标记为ER-α66、ER-α46和ER-α36的泳道代表着用能编码所述的雌激素受体同种型的表达质粒转染、并在转染后2天裂解的HEK 293细胞的分离的培养物。用抗-ER-α抗体(H222),免疫检测每种转染子的裂解物。来自MCF7细胞的细胞提取物用作阳性对照。用箭标指示ER-α66、ER-α46和ER-α36的位置。
图12显示了:(a)能编码ER-α36且包含ER-α36启动子的基因的5′侧翼序列的DNA序列(SEQ ID NO:22),和(b)能编码ER-α36且包含由外显子9编码的核苷酸的基因的3′侧翼序列的DNA序列(SEQ ID NO:25)。在5′侧翼序列中,推定的转录结合位点标有下划线,且也指示了能结合核酸序列的蛋白。也用箭标指示了cDNA的起始位点。
图13显示了不同的乳腺癌细胞MCF10A、T47D、MCF7和MDA-MB-231中的ER-α36的RNA印迹分析。用箭标指示了ER-α36和肌动蛋白的位置。
图14显示了ER-α36对ER-α66和ER-β的AF-1和AF-2结构域介导的转录转反式激活活性的抑制。(+E2),E2处理的细胞;(-E2),未用E2处理的细胞。
图15显示,ER-α36能介导由E2刺激的膜-启动的MAPK激酶途径。(a)蛋白印迹显示了用雌二醇-17β(E2β)处理ER 36-293细胞,会诱导Mek1/2和ERK1/2的快速磷酸化。P-Mek1/2和P-ERK1/2,分别是Mek1/2和ERK1/2的磷酸化形式。(b)血清(而不是E2β)会诱导对照载体-293细胞中的ERK1/2的磷酸化。P-ERK1/2是ERK1/2的磷酸化形式。(c)不同的雌激素和抗雌激素药会诱导ER36-293细胞中的ERK1/2的快速磷酸化。P-ERK1/2是ERK1/2的磷酸化形式。(d)他莫昔芬处理能组成型地刺激ER36-293细胞中的ERK1/2磷酸化。P-ERK1/2是ERK1/2的磷酸化形式。
图16表明,ER-α36能介导E2β诱导的MAPK激酶核信号传导,并刺激细胞生长。(a)E2β对MAPK激酶核信号传导的作用。用5XGal4-LUC,即含有5个Gal4 DNA结合位点的萤光素酶报告质粒,和含有与Gal4DNA结合结构域融合的ELK转录活化结构域的Gal-ELK表达载体(上图),瞬时转染了ER36-293和对照载体-293细胞。转染后,将细胞培养物维持在没有雌激素的培养基中36小时,然后加入E2β(1nM或10nM)12小时。具有标准差的萤光素酶活性代表着一式两份进行的超过3次实验。(b)E2β和抗雌激素药能刺激ER36-293细胞的生长。显示了在490nm的吸光度数据。平均了超过5次独立实验的结果;显示了平均值和SEM。还通过成对的t-检验,评价了这些结果的统计学显著性。ER36-293和载体-293细胞的P-值小于0.001。
图17表明,ER-α36主要是基于膜的雌激素受体。(a)蛋白印迹分析ER-α36在不同的确立的乳腺癌细胞系中的表达。剥离相同的印迹,并用抗-肌动蛋白抗体探测,以确保等量装载。(b)ER-α36在ER-α36转染的293细胞中的亚细胞定位。用ER-α36特异性的抗体,免疫印迹不同的亚细胞级分中的ER-α36。W,全细胞裂解物;PM,质膜;C,胞质溶胶;N,核。通过免疫印迹质膜、胞质溶胶、核和高尔基体的各种蛋白标记,检查亚细胞级分纯度。5′NT,5′核苷酸酶;D4-GDI,GDP解离抑制剂;mSin3A,组蛋白重构复合物的组分;COPB,β外被蛋白。
图18表明,E2β能促进ER-α66阴性的乳腺癌细胞MDA-MB-231在软琼脂中的生长。在不存在E2β(0)、存在10nM E2β(E2)、存在10nM E2β和10nM他莫昔芬(E2+TAM)和存在单独的10nM他莫昔芬(TAM)的情况下,使MDA-MB-231细胞在软琼脂上生长3周。
图19表明,E2β能诱导ER-α66阴性的乳腺癌细胞MDA-MB-231中的膜启动的雌激素信号传导。用雌二醇-17β(E2β)处理MDA-MB-231细胞,会诱导ERK1/2的快速磷酸化。用E2β(10nM)处理细胞不同的时间点,使所述细胞裂解,并使用磷酸化依赖性的和非依赖性的抗体,通过蛋白印迹进行分析。
本发明的优选实施方案的详述
已经发现,小窝蛋白-1(Cav-1)系统的下调会组成型激活促分裂原活化蛋白激酶(MAPK)途径,激活雌激素受体-α(ER-α)的表达,和触发正雌激素信号传导。该发现首次提供了活化的MAPK信号传导和乳房肿瘤发生(尤其是由雌激素刺激的乳腺癌发展)之间的清楚的联系。该发现有力地提示,Cav-1通过协调MAPK和雌激素信号传导途径之间的通讯,在维持乳房上皮细胞的正常生长中起重要作用,且它的下调可能有助于这2个重要途径的调节异常,这最终导致乳腺肿瘤发生。图2显示了雌激素信号传导途径和MAPK信号传导途径的示意图。
还已经鉴别和克隆了雌激素受体-α同种型。从位于原始的66-kDa ER-α(ER-α66)基因的第一个内含子中的启动子,产生了雌激素受体-α的该36-kDa同种型(ER-α36)。ER-α36不同于ER-α66,因为它缺少2个转录活化结构域(AF-1和AF-2),但是保留了DNA-结合、二聚化和大部分的配体-结合结构域。ER-α36的结构表明,ER-α36是雌激素信号传导的调节剂。ER-α36也可以介导雌激素信号传导的膜作用,因为它主要在质膜上表达,且也在胞质溶胶和核中表达。
已经将ER-β视作ER-α66介导的雌激素信号传导的组成型调节剂。发现缺少AF-1结构域的ER-α46可以二聚化成ER-α66,并抑制由ER-α66的AF-1结构域介导的反式激活活性,这表明ER-α46在由ER-α66的AF-1结构域介导的功能活性中起调节作用。ER-α36缺少AF-1和AF-2结构域。因而,认为ER-α36会抑制由ER-α66的AF-1和AF-2介导的生物功能,以及由ER-α46的AF-2介导的功能。利用ER-α36和ER-α46介导的调节,其二者可能在不同的组织中以不同的水平表达,ER-α66可以在不同的靶组织中不同地起作用。认为这样的机理能解释不同生物过程中的雌激素信号传导的多效性作用。
本发明的多肽和肽模仿物(peptidomimetic)
本发明提供了多肽。如本文使用的,术语“多肽”泛指通过肽键连接到一起的2个或更多个氨基酸的聚合物。术语肽、寡肽和蛋白都包含在多肽的定义中,且这些术语可互换地使用。应当理解,这些术语不意味着氨基酸聚合物的特定长度,它们也无意暗示或区分多肽是否由重组技术、化学或酶促合成生成,或是天然产生。本文公开和描述了本发明范围内的多肽的多个实例。在天然产生的多肽或多核苷酸的情况下,优选地,分离这样的多肽或多核苷酸,并任选地纯化。“分离的”多肽或多核苷酸是从它的天然环境分开和分离的多肽或多核苷酸。“纯化的”多肽或多核苷酸是至少60%地不含、优选地75%地不含和最优选地90%地不含它们天然地伴随的其它组分的多肽或多核苷酸。认为在它们天然发生的生物外面生产(例如,通过化学或重组方法)的多肽和核苷酸,是定义的分离的和纯化的,因为它们从未存在于天然环境中。“外源多肽”指外来多肽,即通常不存在于细胞中的多肽,或通常存在于细胞中、但是已经通过实验方法导入细胞中(例如,通过导入能编码该多肽的多核苷酸)的多肽。
本发明的多肽可以是生物活性的。这样的生物活性在本文中称作“ER-α36活性”。可以用于确定本发明的多肽是否是生物活性的生物测定的实例包含,使能表达该多肽的细胞接触雌激素或抗-雌激素药,并测定在有雌激素或抗-雌激素药存在的情况下,与不表达本发明的多肽的对照细胞中的MAPK活性相比,MAPK途径的活性是增高还是降低。优选地,MAPK活性是ERK 1/2和Mek 1/2的磷酸化,且优选地,在有抗-雌激素药存在的情况下,本发明的多肽诱导的ERK1/2的磷酸化不减少。优选地,ER-α36活性是膜启动的。通过暴露能表达具有对不同配体的ER-α36活性的多肽的细胞,可以测量ER-α36活性。可以使用的配体的实例,包括但不限于,雌酮(E1)、17α-雌二醇(E2α)、17β-雌二醇(E2β)、雌三醇(E3)、雌四醇(estetrol)(E4)或附着到不可透过膜的分子,例如牛血清清蛋白(BSA)的雌激素。通常,当待测ER-α36活性限于膜启动的ER-α36活性时,使用附着到不可透过膜的分子上的雌激素。使用的雌激素的量可以变化,且优选地,在1nM至10nM的范围内。暴露于雌激素的细胞优选地是休眠细胞。该暴露允许发生5-90分钟,然后裂解细胞,并通过SDS-聚丙烯酰胺凝胶电泳分离存在于细胞中的多肽。将分离的多肽转移到膜上后,使用针对非磷酸化和磷酸化形式的ERK 1/2和Mek 1/2的抗体,评价MAPK途径的活化。任选地,可以包括抗-雌激素药,例如他莫昔芬、4OH-他莫昔芬或ICI-182,780,以确定ERK 1/2的磷酸化是否对抗雌激素药不敏感。
本发明提供了具有SEQ ID NO:20所述的氨基酸序列的多肽。该多肽,和本文所述的有关多肽,在本文中也称作ER-α36、ER-α36同种型和ER受体α36-亚基。如图1所示,当与ER-α66同种型相比较时,ER-α36同种型缺少氨基-末端氨基酸残基1-183,羧基-末端氨基酸残基430-595,且在它的C-末端具有27个氨基酸残基的添加(见表1)。雌激素受体α异构体包括ER-α36、ER-α46、ER-α66。雌激素受体β异构体包括ER-β。本发明也提供了雌激素受体,其包括ER-α36同种型。无意进行限制,认为ER-α36同种型能通过与ER-α66、ER-α46或ER-β形成二聚体,通过调节雌激素受体功能,来调节细胞对雌激素的反应。而且,认为ER-α36缺少活化因子1(AF-1)和活化因子2(AF-2)活性,并因而缺少内在的转录活性。但是,认为ER-α36保留了完整的二聚化结构域,其允许ER-α36与ER-α46、ER-α66或ER-β二聚化。认为该相互作用允许ER-α36调节含有雌激素受体的ER-α46、ER-α66和ER-β的活性。
表1
氨基酸和核苷酸序列
本发明的多肽包括与SEQ ID NO:20具有至少70%同一性的氨基酸序列的多肽。这样的多肽包括与SEQ ID NO:20具有超过70%的至少单个百分比的同一性的氨基酸序列的多肽,例如与SEQ IDNO:20具有71%、72%、73%的同一性,等等,直到100%的同一性。优选地,多肽包括且有如下氨基酸序列的多肽,所述氨基酸序列以递增的优选等级,与SEQ ID NO:20具有至少约80%同一性、至少约90%同一性或至少约95%同一性。优选地,多肽是生物活性的。优选地,多肽具有36kDa的分子量,如按照十二烷基硫酸钠(SDS)-聚丙烯酰胺凝胶上的电泳测得的。一般,参与ER-α66的磷酸化的残基,例如S236、K302和K303是保守的,参与ER-α66的DNA结合结构域、配体结合结构域和二聚化结构域的功能的那些残基也是如此。在DNA结合、配体结合和/或二聚化中起作用的残基是本领域已知的。
通常,通过比对2个氨基酸序列的残基,确定2个多肽序列之间的百分比同一性,以最优化沿着它们的序列长度的相同氨基酸的数目;在进行比对时允许任一个或2个序列中有缺口,以最优化相同氨基酸的数目,尽管每个序列中的氨基酸必须保持它们的正确次序。优选地,使用BLAST 2搜索算法的Blastp程序,版本2.0.9,如Tatusova等(FEMS Microbiol.Lett.,174,247-250(1999))所述,且可以在http://www.ncbi.nlm.nih.gov/blast/bl2seq/bl2.html从环球网上得到,来对比2个氨基酸序列。优选地,使用所有BLAST 2搜索参数的缺省值,包括矩阵(matrix)=BLOSUM62;开放缺口罚分(open gap penalty)=11,延伸缺口罚分(extension gap penalty)=1,gap x_dropoff=50,期望(expect)=10,字长(wordsize)=3,并任选地,过滤器处于开。在使用BLAST搜索算法对比2个氨基酸序列时,结构相似性称作“同一性”。
本发明也提供了作为ER-α36雌激素受体同种型的片段的多肽。优选地,片段是免疫原性的。在有些方面,片段具有ER-α6活性。免疫原性片段的实例是,SEQ ID NO:1的氨基酸13-27所述的氨基酸序列,更优选地,SEQ ID NO:1的1-27。这样的片段可以用于制备能特异性地结合ER-α36雌激素受体同种型的抗体。片段的实例包括已经在N-末端或C-末端或两末端截短1个或多个氨基酸的雌激素受体同种型,只要该片段含有至少5个邻接氨基酸,更优选地至少7个邻接氨基酸,甚至更优选地至少10个邻接氨基酸,和最优选地至少12个邻接氨基酸。
本发明提供了融合多肽,其具有偶联到本发明的多肽上的载体多肽。载体多肽可以用于增加或降低融合多肽的溶解度。载体多肽也可以用于增加融合多肽的免疫原性,以增加能结合本发明的多肽的抗体的生产。例如,载体多肽可以融合到本发明的多肽片段上,以促进能特异性地结合ER-α36的抗体的生产。这样的片段的实例是具有SEQID NO:1的氨基酸13-27的多肽。本发明不受用于生成本发明的融合多肽的载体多肽类型的限制。载体多肽的实例包括匙孔血蓝蛋白、牛血清清蛋白、卵清蛋白、小鼠血清清蛋白、兔血清清蛋白等。载体多肽也可以用于分离或检测融合多肽。这样的载体蛋白的实例包括谷胱甘肽-S-转移酶、麦芽糖-结合蛋白、壳多糖-结合蛋白和具有下述氨基酸序列的多肽:QFFGLM(SEQ ID NO:2)、EQKLISEEDL(SEQ IDNO:3)、KAEDESS(SEQ ID NO:4)、YPYDVPDYA(SEQ ID NO:5)、DYKDDDDK(SEQ ID NO:6)、YTDIEMNRLGK(SEQ ID NO:7)、MASMTGGQQMG(SEQ ID NO:8)、DTYRYI(SEQ ID NO:9)、TDFYLK(SEQ ID NO:10)、HHHHHH(SEQ ID NO:11)、HPOL(SEQ ID NO:12)、QYPALT(SEQ ID NO:13)、QRQYGDVFKGD(SEQ ID NO:14)、EYMPME(SEQID NO:15)、EFMPME(SEQ ID NO:16)和RYIRS(SEQ ID NO:17)。因此,通过与结合融合多肽的载体多肽部分的其它组分的相互作用,可以检测或分离融合多肽。例如,通过使用已知的方法,可以用生物素检测或分离具有作为载体多肽的抗生物素蛋白的融合多肽。载体多肽也可以用于造成融合多肽在细胞中表达后形成内含体。载体多肽也可以是输出信号,其能造成融合多肽输出细胞,或把融合多肽导向细胞中的隔室,例如周质。
本发明也提供了本发明的2种或更多种多肽,其连续连接进单个氨基酸链中。这样的多肽在本文中称作多肽。多肽可以通过接头连接(见Stahl等,美国专利号6,558,924)。可以分离这样的多蛋白,然后切割,以生成本发明的多肽或偶联多肽。通过使用许多方法,例如化学或蛋白酶切割,可以切割多蛋白。因此,可以将接头设计成被特定的蛋白酶或化学试剂切割。可以用于切割本发明的多蛋白的化合物的实例包括化学试剂和酶。化学试剂的实例包括溴化氰、甲酸和加热、羟胺和加热、溶于醋酸中的亚碘酰基苯甲酸-2-(2-硝基苯基)-3-甲基-3-溴代假吲哚等。酶的实例包括Ala-64枯草杆菌蛋白酶、梭菌蛋白酶、胶原酶、肠激酶、凝血因子Xa、肾素、α-凝血酶、胰蛋白酶、胰凝乳蛋白酶、烟草蚀纹病毒蛋白酶等。多蛋白可以用于增加本发明的多肽的生产效率。生产多蛋白的方法是本领域已知的(见Coolidge等,美国专利号6,127,150)。
本发明的多肽包括已经通过添加、取代或缺失一个或多个邻接的或不邻接的氨基酸来修饰的类似物,或已经被化学地或酶促地修饰的类似物,例如通过报告基团的附着,通过N-末端、C-末端或其它官能团修饰或衍生,或通过环化,只要该类似物能保留生物活性或能刺激结合ER-α36的抗体的生产。类似物因而可以在多肽的一个或两个末端包括其它的氨基酸。优选地,类似物是免疫原性的,更优选地,类似物是免疫原性的,且具有ER-α36活性。在有些方面,本发明提供了非类似物的多肽。
本发明的多肽中的氨基酸取代优选地是保守取代,其选自该氨基酸所属类别的其它成员。例如,蛋白生物化学领域众所周知,属于具有特定大小或特征(例如电荷、疏水性和亲水性)的氨基酸组的氨基酸,通常可以取代为另一种氨基酸,而基本上不改变多肽的结构。例如,非极性的(疏水的)氨基酸包括丙氨酸、亮氨酸、异亮氨酸、缬氨酸、脯氨酸、苯丙氨酸、色氨酸和酪氨酸。极性的中性氨基酸包括甘氨酸、丝氨酸、苏氨酸、半胱氨酸、酪氨酸、天冬酰胺和谷氨酰胺。带正电荷的(碱性)氨基酸包括精氨酸、赖氨酸和组氨酸。带负电荷的(酸性)氨基酸包括天冬氨酸和谷氨酸。优选的保守取代的实例包括Lys取代Arg,反之亦然,以维持正电荷;Glu取代Asp,反之亦然,以维持负电荷;Ser取代Thr,以维持游离的-OH;和Gln取代Asn,以维持游离的NH2。有关的氨基酸(例如3-羟脯氨酸、4-羟脯氨酸、高半胱氨酸、2-氨基己二酸、2-氨基庚二酸、γ-羧基谷氨酸、β-羧基天冬氨酸)、氨基酸酰胺(鸟氨酸、高精氨酸、N-甲基赖氨酸、二甲基赖氨酸、三甲基赖氨酸、2,3-二氨基丙酸、2,4-二氨基丁酸、高精氨酸、肌氨酸和羟赖氨酸)和取代的苯丙氨酸、正亮氨酸、正缬氨酸、2-氨基辛酸、2-氨基庚酸、statine、β-缬氨酸、萘基丙氨酸、四氢异喹啉-3-羧酸和卤化的酪氨酸,可以被交换成类似的氨基酸。
本发明提供了本发明的多肽的肽模仿物。肽模仿物指这样的多肽,其中至少一个肽键已经被替换为非肽键,例如在制药工业中经常用作非肽药物的那些,其具有与模板多肽类似的性质(Fauchere,J.,Adv.Drug Res.,15:29(1986),Evans等,J.Med.Chem.,30:1229(1987),和Janda等,美国专利号6,664,372)。肽模仿物在结构上类似于具有肽键的多肽,但是具有一个或多个通过本领域已知的方法任选地被下述键替代的肽键:例如,--CH2NH--、--CH2S--、--CH2--CH2--、--CH=CH--(顺式和反式)、--COCH2--、--CH(OH)CH2--和--CH2SO--。肽模仿物胜过天然多肽实施方案的优点包括,更经济的生产,更高的化学稳定性,改变的特异性和增强的药理性质,例如半衰期、吸收、效价和功效。
用相同类型的D-氨基酸取代多肽或肽模仿物中的一个或多个氨基酸(例如,D-赖氨酸替代L-赖氨酸),可以产生多肽和肽模仿物,其例如更稳定,且更能抗内源蛋白酶。
通过在氨基-末端和/或羧基-末端添加能减少体内降解的封闭剂,可以修饰本发明的多肽和肽模仿物用于体内用途。这可以用于这样的情形,其中多肽末端倾向于被蛋白酶体内降解。这样的封闭剂包括但不限于,可以附着到本发明的多肽或肽模仿物的氨基和/或羧基末端残基的其它的有关的或无关的肽序列。这可以在化学合成过程中完成,或通过一般技术的人员熟悉的方法,通过重组DNA技术完成。或者,封闭剂例如焦谷氨酸,或本领域已知的其它分子,可以附着到氨基和/或羧基末端残基上,或氨基末端的氨基或羧基末端的羧基可以替换为不同的部分。因此,本发明提供了被封闭的多肽和肽模仿物,和氨基末端、羧基末端或其组合。
通过使用许多表达系统,包括但不限于,用其中已经插入了本发明的多核苷酸的重组载体转化的细胞或微生物,可以小规模或大规模地生产本发明的多肽。下面描述了这样的重组载体和其应用方法。这些载体可以用于转化许多生物。这样的生物的实例包括,细菌(例如,大肠杆菌(E.coli)或枯草芽孢杆菌(B.subtilis));酵母(例如,糖酵母属(Saccharomyces)和毕赤氏酵母属(Pichia));昆虫(例如,杆状病毒);植物;或哺乳动物细胞(例如,COS、CHO、BHK、293、VERO、HeLa、MDCK、W138和NIH 3T3细胞)。直接从用载体转染的哺乳动物得到的原代或次生细胞,也可以用作宿主细胞。
合成方法也可以用于生产本发明的多肽和肽模仿物。这样的方法是本领域已知的和常规的。例如,固相肽合成方法是确定的且广泛使用的方法。通过在免疫亲和或离子交换柱上的分级分离、乙醇沉淀、反相HPLC、硅石或阴离子交换树脂例如DEAE上的色谱、色谱聚焦、SDS-PAGE、硫酸铵沉淀、使用例如Sephadex G-75的凝胶过滤、配体亲和色谱等,可以容易地纯化多肽。通过使融合多肽结合分离介质,然后切割融合多肽以释放纯化的多肽,也可以容易地纯化多肽。例如,可以生成包含在多肽和载体多肽之间的凝血因子Xa切割位点的融合多肽。融合多肽也可以结合亲和柱,后者上面结合了融合多肽的载体多肽部分。然后,可以用凝血因子Xa切割融合多肽,以释放多肽。这样的系统已经与凝血因子Xa去除试剂盒组合使用,用于纯化本发明的多肽。
多核苷酸
本发明提供了能编码本发明的多肽的多核苷酸。术语“多核苷酸”泛指以5′至3′方向共价连接的2个或更多个核苷酸的聚合物。多核苷酸可以包括具有不同功能的核苷酸序列,包括例如编码序列,和非编码序列,例如调节序列。编码序列、非编码序列和调节序列如下面定义。术语核酸、核酸分子和寡核苷酸包含在多核苷酸的定义内,且这些术语可互换地使用。应当理解,这些术语不意味着核苷酸聚合物的特定长度,它们也无意暗示或区分多核苷酸是否由重组技术、化学或酶促合成生成,或是天然产生。
多核苷酸可以是单链的或双链的,且第二个互补链的序列由第一链的序列决定。因此,术语“多核苷酸”应当广义地理解为包括单链的核酸聚合物,它的互补体,和由此形成的双链体。多核苷酸的“互补性”指2个单链多核苷酸彼此碱基配对的能力,其中一个多核苷酸上的腺嘌呤与另一个上的胸苷(或尿嘧啶,在RNA的情况下)碱基配对,而一个多核苷酸上的胞苷与另一个上的鸟嘌呤碱基配对。当一个多核苷酸上的核苷酸序列与第二个多核苷酸上的核苷酸序列碱基配对时,2个多核苷酸彼此互补。例如,5′-ATGC和5′-GCAT是完全互补的,5′-GCTA和5′-TAGC也是如此。
本发明的多核苷酸的一个实例是SEQ ID NO:21(见表1,也即存在于GenBank登记号BX640939中的核苷酸序列的核苷酸234-1166)。优选的本发明的多核苷酸也包括具有与能编码根据本发明的多肽的核苷酸序列“基本上互补的”核苷酸序列的多核苷酸,或这样的核苷酸序列的互补体。“基本上互补的”多核苷酸可以包括至少一个碱基对错配,但是2个多核苷酸仍然具有杂交的能力。例如,2个DNA分子5′-AGCAAATAT和5′-ATATATGCT中的每一个的中间核苷酸不能碱基配对,但是这2个多核苷酸仍然是如本文定义的基本上互补的。如果它们在55℃、2X SSC(SSC:150mM NaCl,15mM柠檬酸三钠,pH7.6)示例的杂交条件下杂交,则2个多核苷酸是基本上互补的。用于本发明目的的基本上互补的多核苷酸优选地共有至少一个至少20个核苷酸长的区域,该共有区域具有至少60%核苷酸同一性、优选地至少80%核苷酸同一性、更优选地至少90%核苷酸同一性和最优选地至少95%核苷酸同一性。特别优选的基本上互补的多核苷酸共有许多这样的区域。优选地,多核苷酸与SEQ ID NO:21具有至少70%同一性的核苷酸序列。更优选地,多核苷酸具有如下核苷酸序列,所述核苷酸序列与SEQ ID NO:21具有超过70%的至少单个百分比的同一性,例如与SEQ ID NO:21具有71%、72%、73%的同一性,等等,直到100%的同一性。甚至更优选地,多核苷酸具有与SEQID NO:21具有至少80%同一性、至少90%同一性或至少95%同一性的核苷酸序列。最优选地,多核苷酸具有与SEQ ID NO:21具有100%同一性的核苷酸序列。与SEQ ID NO:21具有至少70%同一性的多核苷酸具有ER-α36活性。
通常,通过比对2个多核苷酸序列的碱基,确定2个多核苷酸序列之间的百分比同一性,以最优化沿着它们的序列长度的相同碱基的数目;在进行比对时允许任一个或2个序列中有缺口,以最优化相同碱基的数目,尽管每个序列中的碱基必须保持它们的正确次序。优选地,使用BLAST 2搜索算法的Blastn程序,版本2.0.11,也如Tatusova等(FEMS Microbiol.Lett,174,247-250(1999))所述,且可以在http://www.ncbi.nlm.nih.gov/blast/bl2seq/bl2.html从环球网上得到,来对比2个多核苷酸序列。优选地,使用所有BLAST2搜索参数的缺省值,包括匹配奖分(reward for match)=1,错配罚分(penalty for mismatch)=-2,开放缺口罚分=5,延伸缺口罚分=2,gap x_dropoff=50,期望=10,字长=11,并任选地,过滤器处于开。使用CLUSTALW多序列比对软件(J.Thompson等,Nucl.Acids Res.,22:4673-4680(1994)),其可以在www.ebi.ac.uk/clustalw/从环球网上得到,也可以容易地确定2个多核苷酸序列之间的核苷酸序列同一性的位置和水平。
应当理解,能编码本发明的多肽的多核苷酸不限于含有天然产生的基因组或cDNA核苷酸序列的全部或部分的多核苷酸,而是也包括由于遗传密码的简并性,能编码这样的多肽的多核苷酸类别。例如,天然产生的多核苷酸序列SEQ ID NO:21只是能编码具有氨基酸SEQ ID NO:20的多肽的核苷酸序列类别的一个成员。能编码选择的多肽序列的核苷酸序列类别很大但是有限,且参考标准的遗传密码,其中已知不同的核苷酸三联体(密码子)能编码相同的氨基酸,本领域的技术人员能容易地确定类别中的每个成员的核苷酸序列。
能“编码”本发明的多肽的多核苷酸任选地包含编码区和非编码区,且因此应当理解,除非另有相反说明,能“编码”多肽的多核苷酸在结构上不限于能编码多肽的核苷酸序列,而是也包括编码区以外(即,5′或3′)的其它核苷酸序列。“编码区”或“编码序列”是能编码多肽的核苷酸序列,且当置于适当的调节序列的控制下时,其能表达编码的多肽。编码区的边界通常由在它的5′端的翻译起始密码子和在它的3′端的翻译终止密码子决定。“外源编码区”指外来编码区,即通常在细胞中不存在的编码区,或通常存在于细胞中、但是已经通过实验方法导入细胞中、可操作地连接到它通常不可操作地连接的调节区的编码区,或其组合。
本发明的多核苷酸在拓扑学上可以是线状的或环状的。多核苷酸可以是,例如,载体的一部分。载体可以提供进一步克隆(多核苷酸的扩增),即克隆载体,或提供由编码区编码的多肽的表达,即表达载体。载体可以包括,但不限于,质粒、噬菌粒、F因子、病毒、粘粒或噬菌体。载体可以是双链或单链的线状或环状形式。通过整合进细胞基因组或存在于染色体外(例如,作为具有复制起点的自主复制质粒),载体也可以转化原核或真核宿主。载体中的多核苷酸可以在适当的启动子或用于体外转录或在宿主细胞中转录的其它调节序列的控制下,并与其可操作地连接,所述宿主细胞例如真核细胞,或微生物,例如细菌。优选的真核细胞实例包括MDA-MB-231、Hela、CHO和MCF10A细胞系。调节序列,或调节区,指位于编码序列的上游、中间或下游的核苷酸序列,且可操作地连接编码序列。调节序列的实例包括增强子、启动子、翻译前导序列、内含子和多腺苷酸化信号序列。它们包括天然的和合成的序列,以及可以与合成的和天然的序列相组合的序列。调节序列不限于启动子。但是,在本发明中有用的一些合适的调节序列包括,但不限于,组成型启动子、组织-特异性的启动子、发育-特异性的启动子、诱导型启动子和病毒启动子。术语“可操作地连接”指组分的并列,以便它们处于允许它们以目标方式发挥作用的关系。当以在与调节序列相容的条件下实现编码区的表达的方式连接时,调节序列“可操作地连接”到编码区。
载体可以是能在多种宿主中起作用的穿梭载体。载体也可以是克隆载体,其一般含有一个或少数限制性内切核酸酶识别位点,外来DNA序列可以在这里以可确定的方式插入。可以发生这样的插入,而不丧失克隆载体的基本生物功能。克隆载体也可以含有标记基因,其适用于鉴别和选择用克隆载体转化的细胞。标记基因的实例是四环素抗性或氨苄青霉素抗性。许多克隆载体可以商业上得到(例如Stratagene,New England Biolabs,Clonetech)。载体可以是含有调节序列的表达载体,所述调节序列能指导插入表达载体的多核苷酸的表达。许多载体可以商业上得到,且是本领域已知的(Stratagene,La Jolla,CA;New England Biolabs,Beverly,MA)。表达载体可以用于体外转录和翻译测定。
将多核苷酸导入载体的方法,是本领域众所周知的(Sambrook等,Molecular Cloning:A Laboratory Manual,第3版,Cold Spring Harbor Press,Cold Spring Harbor,N.Y.(2001))。简而言之,用一种或多种限制酶(限制性内切核酸酶)处理要插入多核苷酸的载体,生成线性化的载体,其具有平端,具有5′或3′突出端的“粘性”末端,或上面的任意组合。也可以用限制酶处理载体,随后用另一种修饰酶处理,例如聚合酶、外切核酸酶、磷酸酶或激酶,以生成线性化的载体,其具有对将多核苷酸连接进载体有用的特征。用一种或多种限制酶处理要插入载体的多核苷酸,以生成线性化的片段,其具有平端,具有5′或3′突出端的“粘性”末端,或上面的任意组合。也可以用限制酶处理多核苷酸,随后用另一种DNA修饰酶处理。这样的DNA修饰酶包括,但不限于,聚合酶、外切核酸酶、磷酸酶或激酶,以生成多核苷酸,其具有对将多核苷酸连接进载体有用的特征。
然后,根据本领域已知的方法,将处理的载体和多核苷酸连接到一起,以形成含有多核苷酸的构建体(Sambrook等,Molecular Cloning:A Laboratory Manual,第3版,Cold Spring Harbor Press,Cold Spring Harbor,N.Y.(2001))。简而言之,可以在有合适的缓冲液和连接酶存在下,组合处理的核酸片段和处理的载体。然后,在适当的条件下,温育混合物,以允许连接酶将核酸片段连接进载体。
本发明还提供了制备本发明的多肽的方法和制备能编码它们的多核苷酸的方法。方法包括生物的、酶促的和化学的方法,以及其组合,且是本领域众所周知的。例如,使用标准的重组DNA技术,可以在宿主细胞中表达多核苷酸,使用无细胞的基于RNA的系统,可以体外地酶促合成它,或使用化学技术,例如固相肽合成,可以合成它。当使用重组DNA技术时,宿主细胞可以是,例如,细菌细胞、昆虫细胞、酵母细胞或哺乳动物细胞。
本发明也提供了具有启动子活性的多核苷酸。一方面,本发明的启动子包括SEQ ID NO:22所述的核苷酸序列,或其一部分。在另一个方面,本发明的启动子具有如下核苷酸序列,所述核苷酸序列与SEQ ID NO:22具有超过70%的至少单个百分比的同一性,例如与SEQ ID NO:22具有71%、72%、73%的同一性,等等,直到100%的同一性。本文描述了确定百分比同一性的方法。甚至更优选地,启动子具有与SEQ ID NO:22具有至少80%同一性、至少90%同一性、或至少95%同一性的核苷酸序列。本发明的启动子具有雌激素受体(ER)调节的活性。如本文使用的,ER调节的活性指在有ER-α66存在下,优选地,在有能结合雌激素的ER-α66存在下,可操作地连接的编码区的增强表达。本发明的启动子以依赖于和非依赖于雌激素的方式表达。本发明的启动子可以可操作地连接到能编码多肽(包括标记多肽)的编码区。标记多肽的实例包括检测标记(例如,荧光蛋白、酶、抗原标记等)和选择标记(能造成药物抗性、药物易感性或营养缺陷或纠正营养缺陷的多肽等)。
抗体
本发明提供了能特异性地结合本发明的多肽和肽模仿物的抗体。如本文使用的,“能特异性地结合”多肽的抗体是这样的抗体,其仅仅能与诱导该抗体合成的抗原的表位相互作用,或能与结构上有关的表位相互作用。甚至在有多种潜在结合靶存在下,“能特异性地结合”表位的抗体也能在适当的条件下,与表位相互作用。在有些方面,本发明的抗体能特异性地结合ER-α36或其一部分,且不特异性地结合ER-α66或ER-α46。
因此,本发明的多肽和肽模仿物和其片段可以用作抗原,来生产抗体,包括脊椎动物抗体、杂合抗体、嵌合抗体、人源化的抗体、改变的抗体、单价抗体、单克隆和多克隆抗体、Fab蛋白和单结构域抗体。例如,具有SEQ ID NO:1的多肽或其片段,例如SEQ ID NO:1的氨基酸13-27,可以用于产生能特异性地结合ER-α36的抗体。通过将它们共价连接到免疫原性载体上,例如匙孔血蓝蛋白(KLH)、牛血清清蛋白、卵清蛋白、小鼠血清清蛋白、兔血清清蛋白等,可以修饰本发明的多肽或肽模仿物或其片段。
如果需要多克隆抗体,则可以用需要的抗原免疫选择的动物(例如,小鼠、兔子、山羊、马或鸟,例如鸡)。根据已知且常规的方法,收集并处理来自免疫的动物的血清。如果含有针对本发明的多肽的多克隆抗体的血清,含有对其它抗原的抗体,则可以通过免疫亲和色谱,纯化多克隆抗体。本领域已知用于生产和加工多克隆抗血清的技术(见,例如,Harlow等,Antibodies:A Laboratory Manual,Cold Spring Harbor Pub.1988))。
本领域的技术人员也可以容易地生产针对本发明的多肽或肽模仿物或其片段的单克隆抗体。通过杂交瘤制备单克隆抗体的一般方法是众所周知的(见,例如,Harlow等,Antibodies:A Laboratory Manual,Cold Spring Harbor Pub.1988)。通过细胞融合,且也可以通过其它技术,例如用致癌DNA直接转化B淋巴细胞,或用EB病毒转染,可以生成无限增殖化的抗体生产性细胞系(杂交瘤)。可以筛选针对本发明的多肽和肽模仿物生产的单克隆抗体组的各种性质,例如表位亲和性。制备抗体的其它众所周知的方法,包括使用噬菌体显示技术(见,例如,Kay等,Phage display of peptides and proteins:A laboratory manual.San Diego:Academic Press(1996))。
本发明的抗体可以衍生自“人源化的”单克隆抗体。通过将小鼠互补决定区从小鼠免疫球蛋白的重和轻可变链转移进人可变结构域,然后取代鼠对应物的构架区的人残基,可以生产人源化的单克隆抗体。使用源自人源化的单克隆抗体的抗体组分,消除了与鼠恒定区的免疫原性有关的潜在问题。描述了用于克隆鼠免疫球蛋白可变结构域的一般技术(Orlandi等,Proc.Natl Acad.Sci.USA,86:3833(1989),且也描述了生产人源化的单克隆抗体的技术(Jones等,Nature,321:522(1986);Riechmann等,Nature,332:323(1988))。
通过常规的已知方法,包括抗体的蛋白水解,或通过能编码该片段的多核苷酸在大肠杆菌中的表达,可以制备本发明的抗体片段。通过常规方法(用例如胃蛋白酶或木瓜蛋白酶)消化完整的抗体,可以得到抗体片段。例如,通过用胃蛋白酶酶促切割抗体,提供称作F(ab’)2的5S片段,可以生产抗体片段。使用硫醇还原剂和任选的由二硫键的切割产生的巯基的封闭剂,可以进一步切割该片段,以生成3.5SFab′单价片段。或者,使用胃蛋白酶的酶促切割,会直接产生2个单价的Fab′片段和1个Fc片段。
也可以使用切割抗体的其它方法,例如分离重链,以形成单价的轻-重链片段,进一步切割片段,或其它的酶促的、化学的或遗传技术,只要该片段能结合可由完整的抗体识别的抗原。
可以筛选抗体,以确定它们所结合的表位的同一性。表位指抗原(例如本发明的多肽)上的抗体互补位结合的部位。表位通常由分子的化学活性的表面基团例如氨基酸或糖侧链组成,且可以具有特定的三维结构特征,以及特定的电荷特征。可以用于鉴别表位的方法,是本领域已知的(Harlow等,Antibodies:A Laboratory Manual,page319(Cold Spring Harbor Pub.1988)。
可以筛选抗体特异性地结合本发明的多肽或肽模仿物的能力。例如,通过使用本领域的常规方法,可以选择能特异性地结合ER-α36同种型或其部分、但是不能结合ER-α46或α66同种型的抗体(见Kitajima等,美国专利号6,534,281,和Harlow等,Antibodies:ALaboratory Manual,Cold Spring Harbor Pub.1988)。
可以将本发明的抗体偶联到多种化合物上。化合物的实例包括检测标记。这样的检测标记的实例包括荧光标记、酶、放射性同位素等,例如抗生物素蛋白或生物素,其允许检测抗体。本领域已知将抗体偶联到检测标记的方法,和有用的检测标记。这样的抗体可以用于检测ER-α36的自动系统中。抗体可以共价地附着到化疗剂。本领域已知可用于治疗癌症例如乳腺癌和前列腺癌的化疗剂。化疗剂的实例包括苯并二氢吡喃、代马孕酮酯、屈洛西芬、艾多昔芬、他莫昔芬、雷洛昔芬、托瑞米芬、氟维司群(fulvestrant)和faslodex、二膦酸盐、降钙素、tribolone、甲状旁腺素或雷尼酸锶(strontium ranelate)。其它实例包括细胞因子,或毒素,例如白喉毒素A链。
组合物
本发明也提供了组合物,其包含本发明的多核苷酸、肽模仿物或抗体。这样的组合物一般包含药学上可接受的载体。如本文使用的“药学上可接受的载体”包括与药物施用相容的盐水、溶剂、分散介质、涂层、抗细菌剂和抗真菌剂、等渗剂和吸收延缓剂等。也可以将其它活性化合物整合进组合物中。
可以将本发明的组合物制备成许多形式,包括片剂、硬或软胶囊、水溶液、悬浮液和脂质体,和其它缓释制剂,例如成形的聚合凝胶。可以配制经口剂型,从而使多肽、肽模仿物或抗体在经过胃后释放进肠中。这样的制剂记载在Hong等,美国专利号6,306,434和其中包含的文献中。
经口的液体组合物可以是下述形式:例如,水性的或油性的悬浮液、溶液、乳剂、糖浆或酏剂,或可以作为干燥产品存在,以在使用前与水或其它合适的载体配制。这样的液体组合物可以含有常规添加剂,例如悬浮剂、乳化剂、非水载体(其可以包括食用油)或防腐剂。
组合物可以配制成用于肠胃外施用(例如,通过注射,例如,快速浓注或连续输注),且可以以单位剂型存在于安瓿、预装注射器、小体积输注容器或具有添加的防腐剂的多剂量容器中。组合物可以采取下述形式:在油性或水性载体中的悬浮液、溶液或乳剂,且可以含有配制试剂,例如悬浮剂、稳定剂和/或分散剂。
可以将适用于直肠施用的组合物制备成单位剂量栓剂。组合物中可以包含的合适的载体包括盐水溶液示例的那些,和本领域经常使用的其它材料。
对于吸入施用,可以从吹入器、喷雾器或加压包或其它方便的送递气溶胶喷雾剂的工具,方便地送递组合物。加压包可以包含合适的推进剂例如二氯二氟甲烷、三氯氟甲烷、二氯四氟乙烷、二氧化碳或其它合适的气体。在加压的气溶胶的情况下,通过提供阀门来送递计量的量,可以确定剂量单位。
或者,对于通过吸入或吹入施用,组合物可以采取干粉组合物的形式,例如,调节剂和合适的粉末基质(例如乳糖或淀粉)的粉末混合物。粉末组合物可以以单位剂型存在,例如在胶囊或药筒中,或例如,明胶或硬质泡沫塑料衬垫包装,在吸入器或吹入器的辅助下,可以从其中施用粉末。对于鼻内施用,可以通过液体喷雾施用组合物,例如通过塑料瓶喷雾器。
可以将组合物配制成用于经皮施用。也可以将组合物配制成水溶液、悬浮液或分散体、水性凝胶、油包水乳剂或水包油乳剂。通过将组合物被囊化在聚合物中,也可以制备经皮制剂。可以将剂型直接应用于皮肽,作为洗剂、乳膏、油膏,或通过使用贴剂。
本发明的组合物也可以含有其它成分,例如调味剂、着色剂、抗微生物剂和防腐剂。另外,本发明的组合物可以包含药学活性成分,例如激素、抗坏死剂、血管扩张剂、药物试剂等。
通过细胞培养物或实验动物中的标准的药学方法,例如测定LD50(对50%群体致死的剂量)和ED50(对50%群体治疗上有效的剂量),可以确定这样的组合物的毒性和治疗功效。毒性和治疗作用之间的剂量比率,是治疗指数,且其可以表达为比率LD50/ED50。能表现出高治疗指数的化合物是优选的。
从细胞培养测定和动物研究得到的数据,可以用于配制一定范围的人用剂量。这样的化合物的剂量,优选地在循环浓度的范围内,其包括具有较少或无毒性的ED50。依赖于采用的剂型和使用的施用途径,剂量可以在该范围内变化。对于在本发明的方法中使用的化合物,可以从细胞培养测定,初步估计治疗上有效的剂量。可以在动物模型中配制剂量,以达到包含IC50的循环血浆浓度范围(即,能达到症状的半数最大抑制的实验化合物的浓度),如在细胞培养中测定的。这样的信息可以用于更准确地确定在人类中有用的剂量。可以测量血浆中的水平,例如通过高效液相色谱。一方面,人用的剂量范围是足以产生至少10微摩尔(μM,优选地至少25μM,更优选地50μM的血清浓度的量。
可以每天1次或多次至每周1次或多次施用组合物,包括每隔一天1次。熟练的技术人员能明白,某些因素可能影响有效地治疗受试者所需的剂量和时间选择,包括但不限于疾病或障碍的严重性,以前的治疗,受试者的总体健康和/或年龄,和存在的其它疾病。而且,用有效量的组合物治疗受试者,可以包括单一治疗,或优选地,包括系列治疗。
检测方法
本发明提供了检测本发明的多肽的方法。该方法一般包括,提供细胞,分析细胞中的本发明的多肽,和确定细胞是否表达该多肽。细胞可以是离体的或体内的。如本文使用的,术语“离体”指已经从受试者的身体取出(例如,分离)的细胞。离体细胞包括,例如,原代细胞(例如,最近已经从受试者取出,且能在组织培养基中进行有限的生长或维持的细胞)和培养的细胞(例如,能在组织培养基中进行长期生长或维持的细胞)。如本文使用的,术语“体内”指在受试者体内的细胞。体内细胞可以是存在于器官或肿瘤中的细胞。细胞优选地是哺乳动物细胞,例如,小鼠、大鼠或灵长类动物(例如,猴子,人),优选地,人。优选的细胞的实例包括乳房细胞,例如乳腺肿瘤细胞,和前列腺细胞,例如前列腺肿瘤细胞。通过例如,人乳房或前列腺组织的活组织检查,可以从受试者得到细胞。可以使用从几乎所有类型的组织得到的样品。根据本领域已知的方法,可以体外培养对照细胞。不能表达ER-α36kDa雌激素受体并因而可以用作阴性对照的细胞包括HEK293细胞。阳性对照细胞包括,在有血清和BRCA1阴性细胞存在下,可以以低细胞密度生长的细胞。优选地,细胞在有血清存在下以低密度生长。通过例如活组织检查,也可以从组织样品得到对照细胞。
一方面,该方法包括,通过使细胞接触本发明的抗体来分析细胞。使用常规的且本领域已知的检测方法,可以测定细胞是否能表达本发明的多肽。免疫测定的实例包括,竞争性的和非竞争性的测定、例如,放射免疫测定、免疫酶测定、免疫荧光测定或酶免疫测定。也可以使用利用辣根过氧化物酶、碱性磷酸酶或其它化学发光检测剂的化学发光方法。在本发明的方法中,也可以使用蛋白印迹和色谱测定。用于检测本发明的多肽的本发明的抗体可以偶联检测标记,并从而直接检测,或可以使用第二抗体。当使用允许检测不同细胞区域中的多肽的检测方法时,当多肽主要与质膜和细胞质有关、且少于20%信号与核有关时,一般认为能表达ER-α36的细胞是ER-α36阳性的。
在另一个方面,该方法包括,通过扩增多核苷酸,优选地,RNA多核苷酸(例如,mRNA),以形成扩增的多核苷酸,来分析细胞。优选地,通过聚合酶链反应(PCR)扩增多核苷酸,优选地,通过逆转录酶(RT)PCR。从RNA多核苷酸合成DNA多核苷酸的方法,是本领域已知的和常规的。使从细胞得到的多核苷酸接触引物对,后者能扩增多核苷酸,包括SEQ ID NO:22或SEQ ID NO:25,或其组合。由这样的引物对产生的扩增的多核苷酸的存在,指示着细胞能表达雌激素受体。如本文使用的,术语“引物对”指设计用于侧接要扩增的多核苷酸区域的2个寡核苷酸。一个引物与存在于在要扩增的多核苷酸的一端的有义链上的核苷酸互补,而另一个引物与存在于在要扩增的多核苷酸的另一端的反义链上的核苷酸互补。要扩增的多核苷酸可以称作模板多核苷酸。引物与其互补的多核苷酸的核苷酸可以称作靶序列。引物可以具有至少约15个核苷酸,优选地,至少约20个核苷酸,最优选地,至少约25个核苷酸。通过PCR扩增多核苷酸的条件,依赖于使用的引物的核苷酸序列而变化,且用于确定这样的条件的方法,是本领域常规的。
扩增后,可以确定扩增的多核苷酸的存在,例如通过凝胶电泳。通过染色(例如,用溴化乙锭)或用本领域的技术人员已知的合适的标记(包括放射性的和非放射性的标记)进行标记,可以显现扩增的多核苷酸。一般的放射性标记包括33P。非放射性的标记包括,例如,配体例如生物素或洋地黄毒苷,以及酶例如磷酸酶或过氧化物酶,或各种化学发光剂(chemiluminescer)例如萤光素,或荧光化合物如荧光素和它的衍生物。
任选地,也可以测定ER-α66多肽、ER-α46多肽、ER-β或其组合在细胞中的存在。检测ER-α66、ER-α46、ER-β或其组合的存在的方法包括,使用基于抗体或多核苷酸的检测方法,例如多核苷酸的扩增,来应用免疫检测。当使用允许检测不同细胞区域中的多肽的检测方法时,当多肽主要与细胞核有关,例如超过90%的信号与核有关时,一般认为能表达ER-α66或ER-α46的细胞是ER-α66或ER-α46阳性的。
所有乳腺癌中的大约70-80%能表达ER-α66,且称作ER-阳性的乳腺癌。这些肿瘤通常更缓慢地生长,更好地分化,且与更好的总体预后有关(Clark,In:Harris JR,editor.Diseases of the breast,volume 2.Lippincott Williams & Wilkins,38:103-116(2000))。检测本发明的多肽的存在的方法,可以作为诊断标记,用于区分雌激素-阳性的和雌激素-阴性的癌症,优选地,乳腺癌。在本文的实施例中公开的结果强烈表明,由ER-α36介导的雌激素信号传导有助于乳房肿瘤发生,并暗示ER-α36可能参与ER-α66阴性的乳腺癌的肿瘤发生。在本文的实施例中公开的结果也表明,能高度表达本发明的多肽的细胞,比能表达高水平的ER-α66、但是更低水平的ER-α36的细胞,更能耐受低剂量的抗-雌激素药,例如,他莫昔芬。因而,检测本发明的多肽的存在的方法,允许鉴别新的患者类型,即ER-α66-阴性的和ER-α36-阳性的。检测本发明的多肽的存在的方法,也可以用于测定细胞对抗-雌激素药的敏感性,并提供关于例如测定个体的适当的治疗进程的信息。例如,医生可以决定,具有ER-α36-阳性的乳腺肿瘤细胞的受试者,可能需要更高剂量的他莫昔芬,以克服对更低水平的抗性。
检测ER-α66、ER-α46、ER-α36、ER-β或其组合的存在,也允许对比雌激素受体的比率。测定2种或更多种雌激素受体的比率,允许预测细胞对抗-雌激素药(例如,他莫昔芬)的敏感性。例如,在本发明的该方面,测定细胞中ER-α36与ER-α46的比率、ER-α36与ER-α66的比率、ER-α36与ER-β的比率或其组合,并与对照细胞中的对应比率进行对比。这样的比率在本文中称作ER-α36比率。在一个实施例中,对照细胞可以是用抗-雌激素药难以治疗的细胞。在另一个实施例中,对照细胞是用抗-雌激素药不难治疗的细胞。如果在实验细胞中测定的上述ER-α36比率与在对照细胞中所述的ER-α36比率相同,则根据对照细胞的状态,分类实验细胞。例如,如果实验细胞中ER-α36与ER-α66的比率与已知能耐他莫昔芬治疗的对照细胞中ER-α36与ER-α66的比率相同,则将实验细胞归入耐他莫昔芬治疗的。但是,如果实验细胞中ER-α36与ER-α66的比率与不能耐他莫昔芬治疗的对照细胞中ER-α36与ER-α66的比率相同,则将实验细胞归入不耐他莫昔芬治疗的。在另一个实例中,将在实验细胞中测定的ER-α36比率与已知能耐他莫昔芬治疗的对照细胞中的对应比率进行对比,并与已知不能耐他莫昔芬治疗的对照细胞中的比率进行对比。然后如上所述,将实验细胞归入耐他莫昔芬治疗的,或不耐他莫昔芬治疗的。已知不耐他莫昔芬治疗的对照细胞的一个实例是MCF7(ATTC保藏登记号HTB-22)。已知耐低剂量他莫昔芬治疗的对照细胞的一个实例是MDA-MB-231。通过对比实验细胞中的ER-α36比率,也可以将已知耐他莫昔芬治疗的乳腺癌细胞用作对照细胞。
鉴别能结合本发明的多肽的试剂
本发明也提供了鉴别能结合本发明的多肽的试剂的方法。这样的方法也称作筛选测定。该方法包括,组合本发明的多肽和试剂,和测定该试剂是否结合多肽。一般,测定试剂是否结合多肽,包括检测试剂和多肽之间的复合物形成。测定复合物的方法包括,例如,直接检测试剂与多肽的结合,和使用竞争结合测定,检测试剂与多肽的结合。测定可以是无细胞的测定。可以在有或没有雌激素或抗-雌激素药存在下,进行测定。任选地,该方法也包括,测定试剂是否结合ER-α66多肽,例如具有SEQ ID NO:18所述的氨基酸序列的多肽。优选地,试剂不能结合ER-α66多肽。
使用本领域已知的组合文库方法中的许多方法的任一种,包括生物学文库,空间上可寻址的平行固相或液相文库,需要解卷积的合成文库方法,“一个珠子一种化合物(one-bead one-compound)”文库方法,和使用亲和色谱选择的合成文库方法,可以得到试剂。生物文库方法包括肽文库,而其它4种方法适用于肽、非肽寡聚物或小分子化合物文库(Lam,Anticancer Drug Des.12:145(1997))。在本领域可以找到关于合成分子文库的方法的实例(见,例如DeWitt等Proc.Natl.Acad.Sci.USA 90:6909(1993);Erb等.Proc.Natl.Acad.Sci.USA 91:11422(1994);Zuckermann等J.Med.Chem.37:2678(1994);Cho等Science 261:1303(1993);Carrell等Angew.Chem.Int.Ed.Engl.33:2059(1994);Carell等Angew.Chem.Int.Ed.Engl.33:2061(1994);和Gal lop等J.Med.Chem.37:1233(1994))。要筛选的潜在试剂的来源包括,例如,细菌和真菌的发酵培养基,和植物和其它增殖体的细胞提取物。
化合物文库可以存在于例如溶液中(例如Houghten Bio/Techniques 13:412-421(1992)),或珠子上(Lam Nature 354:82-84(1991)),芯片上(Fodor Nature 364:555-556(1993)),细菌(美国专利号5,223,409),孢子(美国专利号5,571,698;5,403,484;和5,223,409),质粒(Cull等Proc.Natl.Acad.Sci.USA 89:1865-1869(1992)),或噬菌体(Scott等Science 249:386-390(1990);Devlin Science 249:404-406(1990);Cwirla等Proc.Natl.Acad.Sci.USA 87:6378-6382(1990);和Felici J.Mol.Biol.222:301-310(1991))。
通过例如用放射性同位素或酶标记偶联试剂,从而可以通过检测复合物中的标记的化合物,测定试剂与本发明的多肽的结合,来确定试剂结合本发明的多肽的能力。例如,可以用125I、35S、14C或3H直接地或间接地标记试剂,并通过直接计数radioemmission或通过闪烁计数,检测放射性同位素。或者,可以用例如,辣根过氧化物酶、碱性磷酸酶或萤光素酶,酶促地标记试剂,并通过测定适当的底物向产物的转化,检测酶标记。
以类似的方式,人们可以测定试剂改变(例如,刺激或抑制)本发明的多肽与多肽的已知配体(例如,本发明的多肽能天然地结合或与其相互作用的分子)的结合的能力。这样的配体的实例是雌激素,以及抗-雌激素药,例如他莫昔芬。在一个优
雌激素受体和使用方法专利购买费用说明
![]()
Q:办理专利转让的流程及所需资料
A:专利权人变更需要办理著录项目变更手续,有代理机构的,变更手续应当由代理机构办理。
1:专利变更应当使用专利局统一制作的“著录项目变更申报书”提出。
2:按规定缴纳著录项目变更手续费。
3:同时提交相关证明文件原件。
4:专利权转移的,变更后的专利权人委托新专利代理机构的,应当提交变更后的全体专利申请人签字或者盖章的委托书。
Q:专利著录项目变更费用如何缴交
A:(1)直接到国家知识产权局受理大厅收费窗口缴纳,(2)通过代办处缴纳,(3)通过邮局或者银行汇款,更多缴纳方式
Q:专利转让变更,多久能出结果
A:著录项目变更请求书递交后,一般1-2个月左右就会收到通知,国家知识产权局会下达《转让手续合格通知书》。









动态评分
0.0