






专利摘要
本发明涉及生物合成工程与微生物制造技术领域,提供了一种异源合成广霍香醇的方法和微生物。具体地,本发明提供了微生物异源合成倍半萜广藿香醇的方法,主要包括对广藿香醇合成酶的密码子优化,合成下游途径的组装与表达,微生物宿主萜类合成前体IPP/DMAPP的强化,最终实现广藿香醇在微生物体内的高效合成。
权利要求
1.一种基因组合,其特征在于,所述的组合包括(i)pts基因的多核苷酸序列和(ii)ispA基因的多核苷酸序列。
2.如权利要求1所述的基因组合,其特征在于,所述组合还包括选自下组的一种或多种多核苷酸序列:
(a)dxs2基因的多核苷酸序列;
(b)idi基因的多核苷酸序列;
(c)MVA途径的合成蛋白基因的多核苷酸序列,较佳地所述合成蛋白基因选自:atoB、mvaS、mvaE,mvaK1、mvaK2、mvaD、或其组合。
3.如权利要求1所述的基因组合,其特征在于,其特征在于,所述的基因组合包括:
第一序列,其中所述第一序列含有pts基因的多核苷酸序列;以及
第二序列,其中所述第二序列含有ispA基因的多核苷酸序列。
4.一种构建物,其特征在于,所述的构建物含有第一表达盒和第二表达盒,其中所述第一表达盒表达pts蛋白;而所述的第二表达盒表达ispA蛋白。
5.一种宿主细胞,其特征在于,所述的宿主细胞中含有携带外源的第一表达盒的第一表达载体和外源的第二表达盒的第二表达载体,或者所述的宿主细胞的基因组中整合有外源的第一表达盒和外源的第二表达盒,
其中所述第一表达盒表达pts蛋白;而所述的第二表达盒表达ispA蛋白。
6.如权利要求5所述的宿主细胞,其特征在于,所述的宿主细胞包括原核细胞和真核细胞。
7.一种生产广藿香醇的方法,其特征在于,包括步骤:
培养权利要求5所述的宿主细胞,从而表达pts蛋白和ispA蛋白,并获得含广藿香醇的发酵产物;和
从所述发酵产物中分离和/或纯化所述的广藿香醇。
8.一种编码广藿香醇合成酶的核苷酸序列,其特征在于,所述的核苷酸序列编码SEQIDNO:2所示的氨基酸序列,而且所述核苷酸序列的的编码区与SEQIDNO:3所示核苷酸序列有95%以上相同性。
9.如权利要求8所述的核苷酸序列,其特征在于,该核苷酸序列如SEQIDNO:3所示。
10.权利要求1所述的基因组合、权利要求4所述的构建物、或权利要求5所述的宿主细胞的用途,其特征在于,用于在植物体外合成广藿香醇和/或其衍生物。
说明书
技术领域
本发明涉及生物合成工程与生物制造技术领域,具体地提供了一种异源合成广霍香醇的方法和微生物。
背景技术
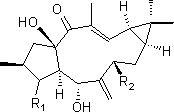
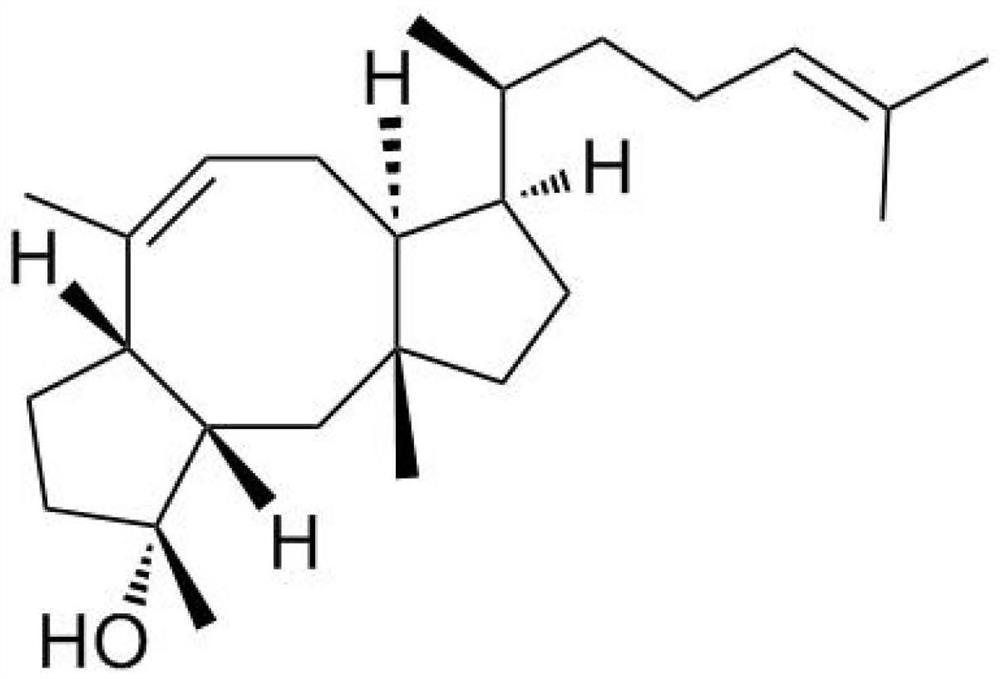
广藿香醇(Patchoulialcohol)是一种重要的三环倍半萜类化合物。广藿香醇又称百秋李醇,其分子式为C15H26O,分子量为222.37,结构如图1所示。
天然形式的广霍香醇主要存在于唇形科刺蕊草属植物广藿香(Pogostemoncablin(Blanco)Benth)的挥发油—广藿香油中,是广藿香挥发油的主要成分,是历版《中国药典》规定的评价广藿香药材及广藿香油质量标准的指标性成分。
广藿香醇单体为无色晶体,有较淡的广藿香香气,其熔点为55-56℃,沸点280℃(常压下),相对密度为1.0284,旋光度-97.4°(c=24,氯仿);不溶于水,溶于醇、醚和常用有机溶剂。
广藿香醇具有独特持久的香味,作为香料的标志性成分被广泛用于日用品和化妆品中。
近年来,随着科研人员对广藿香醇研究的深入,广藿香醇被发现具有很好的神经保护、抗流感病毒、抗多种真菌、抗肿瘤、抗炎症、抗锥虫病原体和毒杀白蚁等生物活性。此外,现代药理实验表明,广藿香醇还具有抗氧化等药理活性,有一定的新药开发价值。
与大多数植物源天然产物一样,目前广藿香醇主要通过化学合成和有机溶剂萃取两种方法制备。
作为含有5个手性碳原子的四面体结构,广藿香醇的化学合成方法过于复杂,且得率极低。
有机溶剂萃取法主要通过天然的含广霍香醇的植物进行组织抽提来获取广霍香醇,然而该方法受到植物生长缓慢、广霍香醇含量低、需要消耗有机溶剂等影响。
JoeChappell等通过转基因的方法对烟草进行遗传改造,使得烟草能够合成0.5μg/g湿重的广藿香醇。然而,植物基因工程方法仍具有植物生长缓慢、广霍香醇含量低、需要消耗有机溶剂等缺点。
由于广藿香醇具有神经保护、抗流感病毒等作用,并且,广藿香醇单体在国外日化产品行业有较成熟的应用基础,而在国内日化产品市场应用仍接近空白,因此,其市场空间广泛,但亟需可解决其高成本难题的工业化生产方法。
虽然人们尝试开发在微生物中生产广霍香醇的技术,然而迄今为止,尚未有在微生物中成功地合成广藿香醇的报道。此外,微生物发酵最大的瓶颈在于如何实现工业化,而工业化的必要条件是其产量需要满足可以工业化的水平。
综上所述,本领域迫切需要开发新的高效、简便、低成本地生产广霍香醇的方法,尤其是通过微生物制备纯度较高的广霍香醇的新方法。
发明内容
本发明的目的就是提供通过微生物生产广霍香醇的新工艺,实现广藿香醇更清洁,更经济的制造。具体来说,本发明的目的是提供一种微生物异源合成倍半萜广藿香醇的的方法。
在本发明的第一方面,提供了一种基因组合,所述的组合包括(i)pts基因的多核苷酸序列和(ii)ispA基因的多核苷酸序列。
在另一优选例中,所述组合还包括选自下组的一种或多种多核苷酸序列:
(a)dxs2基因的多核苷酸序列;
(b)idi基因的多核苷酸序列;
(c)MVA途径的合成蛋白基因的多核苷酸序列,较佳地所述合成蛋白基因选自:atoB、mvaS、mvaE,mvaK1、mvaK2、mvaD、或其组合。
在另一优选例中,所述的组合由(i)pts基因和(ii)ispA基因二种基因构成。
在另一优选例中,所述的基因组合用于在植物体外合成广藿香醇和/或其衍生物。
在另一优选例中,所述的基因组合为多核苷酸序列和/或载体的组合。
在另一优选例中,所述的基因组合包括:
第一序列,其中所述第一序列含有pts基因的多核苷酸序列;以及
第二序列,其中所述第二序列含有ispA基因的多核苷酸序列。
在另一优选例中,所述的基因组合包括:
第一载体,其中所述第一载体含有pts基因的多核苷酸序列;以及
第二载体,其中所述第二载体含有ispA基因的多核苷酸序列。
在另一优选例中,所述的第一载体和第二载体是同一载体(即pts基因的多核苷酸序列和ispA基因的多核苷酸序列位于同一载体上)。
在另一优选例中,所述的基因的多核苷酸序列包括基因组序列和cDNA序列。
在本发明的第二方面,提供了一种构建物,所述的构建物含有第一表达盒和第二表达盒,其中所述第一表达盒表达pts蛋白;而所述的第二表达盒表达ispA蛋白。
在另一优选例中,所述的构建物用于在植物体外合成广藿香醇和/或其衍生物。
在另一优选例中,所述的构建物为表达载体。
在另一优选例中,所述的构建物为1个或2个表达载体。
在另一优选例中,第一表达盒从5’至3’依次含有启动子、pts蛋白编码序列、终止密码子和任选的3’-UTR。
在另一优选例中,第二表达盒从5’至3’依次含有启动子、ispA蛋白编码序列、终止密码子和任选的3’-UTR。
在另一优选例中,所述的pts蛋白编码序列是序列优化的编码序列,较佳地如SEQIDNO.:3所示。
在另一优选例中,所述的ispA蛋白编码序列是序列优化的编码序列,较佳地如SEQIDNO.:6所示。
在本发明的第三方面,提供了一种宿主细胞,所述的宿主细胞中含有携带外源的第一表达盒的第一表达载体和外源的第二表达盒的第二表达载体,或者所述的宿主细胞的基因组中整合有外源的第一表达盒和外源的第二表达盒,
其中所述第一表达盒表达pts蛋白;而所述的第二表达盒表达ispA蛋白。
在另一优选例中,所述的第一载体和第二载体是同一载体(即pts基因的多核苷酸序列和ispA基因的多核苷酸序列位于同一载体上)。
在另一优选例中,所述的宿主细胞包括原核细胞和真核细胞。
在另一优选例中,所述的真核细胞包括酵母。
在另一优选例中,所述的真核细胞是除了广藿香(Pogostemoncablin)之外的植物细胞。
在另一优选例中,所述的原核细胞是大肠杆菌。
在另一优选例中,所述的宿主细胞选自下组:酿酒酵母、大肠杆菌、枯草芽胞杆菌。
在另一优选例中,所述的宿主细胞还含有第三表达载体,所述的第三表达载体携带用于表达选自下组蛋白的第三表达盒:
(a)dxs2蛋白;
(b)idi蛋白;
(c)MVA途径的合成蛋白,较佳地选自:atoB、mvaS、mvaE,mvaK1、mvaK2、mvaD、或其组合;
(d)上述(a)、(b)、(c)的任意组合。
在另一优选例中,所述的(c)是atoB、mvaS、mvaE,mvaK1、mvaK2、mvaD六种蛋白所构成的组合。
在另一优选例中,所述的第三表达盒同时表达dxs2蛋白和idi蛋白。
在本发明的第四方面,提供了一种生产广藿香醇的方法,包括步骤:
培养本发明第三方面所提供的宿主细胞,从而表达pts蛋白和ispA蛋白,并获得含广藿香醇的发酵产物;和
从所述发酵产物中分离和/或纯化所述的广藿香醇。
在本发明的第五方面,提供了一种编码广藿香醇合成酶的核苷酸序列,所述的核苷酸序列编码SEQIDNO:2所示的氨基酸序列,而且所述核苷酸序列的的编码区与SEQIDNO:3所示核苷酸序列有95%以上相同性。
在另一优选例中,该核苷酸序列如SEQIDNO:3所示。
在本发明的第六方面,提供了本发明第一方面所提供的基因组合、本发明第二方面所提供的构建物、或本发明第三方面所提供的宿主细胞的用途,用于在植物体外合成广藿香醇和/或其衍生物。
应理解,在本发明范围内中,本发明的上述各技术特征和在下文(如实施例)中具体描述的各技术特征之间都可以互相组合,从而构成新的或优选的技术方案。限于篇幅,在此不再一一赘述。
附图说明
图1显示了倍半萜广藿香醇的结构式。
图2显示了质粒pJF63构建示意图。
图3显示了质粒pJF678图谱。
图4显示了质粒pJF680图谱。
图5显示了工程菌株BL21(DE3)pJF707的GC检测图谱。
图6显示了工程菌株BL21(DE3)pJF707产广藿香醇的质谱图。
图7显示了构建后各菌株的广藿香醇的产量(mg/L)。
具体实施方式
广藿香醇由萜类化合物的通用前体焦磷酸异戊烯酯(IPP)和二甲烯丙基焦磷酸(DMAPP)经法尼基焦磷酸合酶和广藿香醇合成酶两步反应合成。发明人经过广泛而深入的研究,出乎意料地发现将pts基因和ispA基因作为基础基因组合,以此通过对植物来源的广藿香醇合成酶基因进行密码子优化,构建适于微生物系统表达的下游产物合成途径,实现首次体外合成广藿香醇单体。此外,通过强化MEP途径和引入MVA途径,可进一步实现广藿香醇的异源高效合成,在此基础上完成了本发明。
pts基因
如本文所用,pts基因的全称为广藿香醇合成酶基因(patchoulolsynthase基因),编码广藿香醇合成酶PTS,其NCBI登录号为AY508730,序列如SEQIDNO.:1所示,其中ORF位于第1-1659位,编码长度为552个氨基酸的PTS蛋白(SEQIDNO.:2)。一种特别优化的、适合大肠杆菌表达的pts基因如SEQIDNO.:3所示。
ispA基因
如本文所用,ispA基因的全称为法尼基焦磷酸合成酶基因(farnesylpyrophosphatesynthase基因),编码法尼基焦磷酸合成酶FPPS,其NCBI登录号为U00096,序列如SEQIDNO.:4所示,其中ORF位于第1-900位,编码长度为299个氨基酸的ispA蛋白(SEQIDNO.:5)。
MEP途径的合成酶
类异戊二烯类物质在生物体的生长发育过程中起着很重要的作用。在植物中,这类物质的合成是通过两条途径来完成的:(1)定位于细胞质中的MVA途径;(2)定位于质体中的MEP途径。
其中,MEP途径即甲基苏糖醇磷酸酯途径,亦称作Rohmer途径,主要参与二萜、单萜、类胡萝卜素、异戊二烯等的生物合成。
MEP途径以丙酮酸和3-磷酸甘油醛为前体,经8步反应后合成IPP/DMAPP前体。其中第一步反应1-脱氧木酮糖-5-磷酸合成酶(1-deoxy-D-xylulose5-phosphate,DXS)是MEP途径第一步限速步骤。不同于大肠杆菌,阿维链霉菌含有丰富的萜类次级代谢,其MEP途径含有dxs1,dxs2两个不同拷贝的dxs基因。
此外异戊烯焦磷酸异构酶(Isopentyldiphosphate,IDI)负责DMAPP和IPP的异构化反应。idi基因的表达水平也是影响萜类合成的重要限速步骤。
MVA途径的合成酶
MVA途径主要合成的类异戊二烯类物质为固醇,倍半萜,泛醌和多萜等。MVA途径的催化乙酰-CoA合成IPP/DMAPP,依次涉及乙酰乙酰-CoA合成酶(acetoacetyl-CoA,ACOT),经3-羟基-3-甲基戊二酰-CoA合成酶(hydroxymethylglutaryl-CoAsynthase,HMGS)的催化形成3-羟基-3-甲基戊二酰-CoA合成酶(3-hydroxy-3-methylglutarylcoenzymeA,HMGCoA),再经3-羟基-3-甲基戊二酰-CoA合成酶(hydroxymethylglutaryl-CoAreductase,HMGR)的催化下生成甲羟戊酸(mevalonate,MVA),之后在MVA激酶(mevalonatekinase,MK)的作用下形成磷酸甲羟戊酸(mevalonate-5-phosphate,MVAP),接着在磷酸甲羟戊酸激酶(phosphomevalonatekinase,PMK)的作用下生成焦磷酸甲羟戊酸(mevalonate-5-diphosphate,MVAPP),最终在甲羟戊酸焦磷脱羧酶(mevalonatepyrophosphatedecarboxylase,MPD)的作用下生成萜类物质合成前体异戊烯焦磷酸(Isopenteny,IPP)。大肠杆菌中自身仅以MEP途径供应萜类前体,仅存在着atoB基因编码MVA途径的第一个酶ACOT。不同于大肠杆菌,粪肠球菌和金黄色葡萄球菌等革兰氏阳性菌以MVA途径作为萜类前体合成途径。基于本研究组前期的工作,本发明人选取了最佳的MVA途径基因模块即粪肠球菌中编码HMGS的基因mvaS,编码HMGR的基因mvaE,以及金黄色葡萄球菌中编码MK的基因mvaK1,编码PMK基因的mvaK2,编码MPD基因mvaD,组装MVA途径。
表达载体、宿主细胞和重组方法
本发明还提供了基于本发明的上述基础基因组合和优选基因组合的构建物、表达盒和载体等。
在本发明中,一种代表性的基因表达盒从5’-3’依次具有下列组件:本发明的启动子、外源基因ORF序列和终止子。
本发明还提供了一种重组载体,其包含本发明所述广藿香醇基础基因组合(或优选基因组合)的表达盒。在优选的实施方式中,所述重组载体的启动子下游包含多克隆位点或至少一个酶切位点。需要表达目的基因时,将目的基因连接入适合的多克隆位点或酶切位点,从而可操作地连接目的基因与启动子。
在另一优选的实施方式中,所述重组载体在5’到3’方向上包括:启动子,目的基因和终止子。
如果需要,所述重组载体还可以包括以下组件:3’多聚核苷酸化信号;非翻译核酸序列;转运和靶向核酸序列;抗性选择标记(二氢叶酸还原酶、新霉素抗性、潮霉素抗性以及绿色荧光蛋白等);增强子;或操作子。
用于制备重组载体的方法是本领域普通技术人员所熟知的。表达载体可以是细菌质粒、噬菌体、酵母质粒、植物细胞病毒、哺乳动物细胞病毒或其它载体。总之,只要其能够在宿主体内复制和稳定,任何质粒和载体都是可以被采用的。
本领域普通技术人员可以采用熟知的方法构建含有本发明所述广藿香醇基础基因组合(或优选基因组合)的表达载体。这些方法包括体外重组DNA技术、DNA合成技术、体内重组技术等。
本发明的表达盒或载体,可以用于转化适当的宿主细胞,以使宿主表达蛋白质。
此外,表达载体优选地包含一个或多个选择性标记基因,以提供用于选择转化的宿主细胞的表型性状,如二氢叶酸还原酶、新霉素抗性、潮霉素抗性以及绿色荧光蛋白(GFP)等。包含上述适当的启动子和目的基因的载体,可以用于转化适当的宿主细胞,以使其能够表达蛋白质。
用于制备重组载体的方法是本领域普通技术人员所熟知的。表达载体可以是细菌质粒、噬菌体、酵母质粒、植物细胞病毒、哺乳动物细胞病毒或其它载体。总之,只要其能够在宿主体内复制和稳定,任何质粒和载体都是可以被采用的。
本发明的表达盒或载体,可以用于转化适当的宿主细胞,以使宿主表达蛋白质。宿主细胞可以是原核细胞,如大肠杆菌,链霉菌属、农杆菌:或是低等真核细胞,如酵母细胞;或是高等真核细胞,如植物细胞。本领域一般技术人员都清楚如何选择适当的载体和宿主细胞。
特别优选的宿主细胞是原核细胞(如大肠杆菌)和低等真核细胞(如酵母)。
用重组DNA转化宿主细胞可用本领域技术人员熟知的常规技术进行。当宿主为原核生物(如大肠杆菌)时,可以用CaCl2法处理,也可用电穿孔法进行。当宿主是真核生物,可选用如下的DNA转染方法:磷酸钙共沉淀法,常规机械方法(如显微注射、电穿孔、脂质体包装等)。
另一类优选的宿主细胞是植物细胞。此外,本发明还提供了用所述转化的植物细胞所再生的植物(尤其是除了广藿香植物的其它植物,如烟草、水稻、小麦、棉花等)。
转化植物也可使用农杆菌转化或基因枪转化等方法,例如叶盘法、幼胚转化法、花芽浸泡法等。对于转化的植物细胞、组织或器官可以用常规方法再生成植株,从而获得转基因的植物。
所述的多核苷酸在高等真核细胞中表达时,如果在载体中插入增强子序列时将会使转录得到增强。增强子是DNA的顺式作用因子,通常大约有10到300个碱基对,作用于启动子以增强基因的转录。本领域一般技术人员都清楚如何选择适当的载体、启动子、增强子和宿主细胞。用重组DNA转化宿主细胞可用本领域技术人员熟知的常规技术进行。
本发明的主要优点包括:
(1)在原核生物中成功实现广藿香醇的高效异源合成。
(2)本发明利用微生物实现了广藿香醇的合成,该技术可适用于广藿香醇的微生物大规模发酵生产。
(3)本发明提供了可生物合成广藿香醇的微生物工程菌株,完成了对广藿香醇合成酶的酶学特性改造,可应用于工业规模提高底物特异性等技术领域。
(4)本发明利用了其它微生物来源的MEP途径基因强化MEP途径,再此基础上引入了不同来源MVA途径基因装配的MVA途径,用于萜类前体的强化,成功地提高了广藿香醇的合成。
下面结合具体实施例,进一步阐述本发明。应理解,这些实施例仅用于说明本发明而不用于限制本发明的范围。下列实施例中未注明具体条件的实验方法,通常按照常规条件,例如Sambrook等人,分子克隆:实验室手册(NewYork:ColdSpringHarborLaboratoryPress,1989)中所述的条件,或按照制造厂商所建议的条件。除非另外说明,否则百分比和份数是重量百分比和重量份数。
通用方法和材料
工程菌株的发酵
将相关的质粒组合共转化常规的大肠杆菌BL21(DE3),挑取单菌落过夜培养12h,离心收集菌体后,加入等体积15%的甘油作为摇瓶发酵的种子,-80℃冻存。按2%的接种量,将冻存的种子液接种到含10mlTB培养基的100ml摇瓶中,同时20%的十二烷和终浓度为0.1mM的IPTG,至于28℃,250rpm的摇床上培养。
广藿香醇的定量检测
采用ThermoScientificTRACEGC-ISQGC-MS系统对广藿香醇进行分析。取十二烷相与水相混匀状态下的发酵液1ml,加入1ml乙酸乙酯,边涡旋边萃取20min。然后离心收集有机相进行GC-MS分析。GC-MS的分析条件为:HP-5毛细管柱(30m×250μM×0.25μM),进样量1μl,不分流,进样温度250℃,柱子初始温度140℃,维持0.5min,以4℃/min,升温至230℃维持3min。将产物的MS谱图与99%纯度的广藿香醇MS谱图对照,鉴定产物峰。同时用广藿香醇标准品绘制标准曲线Y=(8.40E+07)X+(1.45E+07)(Y表示MS峰面积,X表示广藿香醇浓度(mg/L),线性范围为1~40mg/L,R2=0.999)。
质粒:
表1为本发明涉及的质粒;
表2为本发明涉及的PCR引物。
表1
表2
缩写:EC为E.coli,SAV为S.avermitilis,BS为B.subtilis,SAU为Staphylococcusaureus,EF为Enterococcusfaecalis。下划线表示克隆所用酶切位点;粗体表示SpeI酶切位点;斜体表示pET21c载体XbaI和NdeI酶切位点间含核糖体结合位点RBS的间隔序列。
在实施例1-5中,以E.coli作为微生物宿主进行微生物异源合成倍半萜广藿香醇。
实施例1
广藿香醇合成酶基因密码子优化及其人工合成:
为了改善植物来源基因在微生物细胞中的表达,首先需要根据不同宿主细胞密码子偏向性对基因序列进行优化。本发明人选取广藿香的广藿香醇合成酶基因序列(pts,NCBI登录号AY508730),
本实施例中,以E.coli宿主作为实例,按E.coli密码子偏好性等多种因素,进行基因序列的优化,并在优化序列的终止密码子“TAA”后加上SpeI位点序列“ACTAGT”。人工合成后的完整基因如SEQIDNO.:3所示,其中ORF位于1-1659位。
委托由上海捷瑞生物工程有限公司负责合成优化后的pts基因。
实施例2
广藿香醇下游合成途径的构建:
为了赋予微生物细胞能够合成异源天然产物的能力,首先需要组装适合不同微生物宿主异源表达的广藿香醇的合成途径。在植物广藿香中,广藿香醇由萜类的通用前体焦磷酸异戊烯酯(IPP)和二甲烯丙基焦磷酸(DMAPP)经法尼基焦磷酸合酶和广藿香醇合成酶两步反应合成。
在本实施例中,为了在微生物宿主实现广藿香醇的生产,对微生物内源的法尼基焦磷酸合酶基因过表达(也可引入其它生物来源的法尼基焦磷酸合酶基因)。以E.coli宿主为例,为了构建高效的异源途径,本发明人采用T7启动子进行广藿香醇合成酶的表达,同时过表达了E.coli的法尼基焦磷酸合酶基因(ispA,其为野生型),构建非天然的多顺反子表达单元T7Prom-pts-ispA-T7Term。
将人工合成的pts基因克隆到了pET28a(购自EMD4Biosciences)的NcoI和HindIII位点上,获得质粒pJF706。
通过常规PCR获得ispA基因的完整ORF序列(SEQIDNO.:4)的扩增产物,该扩增产物的两端分别带有NcoI和EcoRI位点。将所述扩增产物经NcoI和EcoRI双酶切后,插入市售的pET28a为载体NcoI和EcoRI位点,从而得到pJF21。
用XbaI和HindIII双酶切质粒pJF21,胶回收含ispA基因的DNA片段,然后将其连接到质粒pJF706的SpeI和HindIII位点,构建质粒pJF707。
将含异源合成途径的质粒pJF707导入不同系列的E.coli宿主中,即可获得能够合成广藿香醇的重组菌株。以E.coliBL21(DE3)菌株为例,转化后获得重组大肠杆菌BL21(DE3)pJF707。
按上述的种子制备和摇瓶发酵条件对菌株进行发酵培养。按上述的GC-MS检测条件对发酵产物进行成分分析。
结果表明:重组菌株大肠杆菌BL21(DE3)pJF707成功地合成了广藿香醇(如图5,图6),广藿香醇的产量为4.65mg/L。
实施例3
萜类合成前体供应的强化:
为了增强广藿香醇在重组微生物中的合成,需要对宿主的萜类前体合成途径进行强化,增强IPP/DMAPP的供应。在本实施例中,首先引入强化了大肠杆菌自身MEP途径关键基因dxs(EC)和idi(EC),构建了用于共表达的dxs(EC)和idi(EC)质粒和菌株。
以大肠杆菌基因组DNA作为模板,用引物(dxs(EC)F和dxs(EC)R,idi(EC)F和idi(EC)R)分别PCR扩增dxs(EC)和idi(EC)。将纯化后的PCR产物dxs(EC)和idi(EC)分别用NcoI和EcoRI双酶切后,连接到pET21d的相应酶切位点间分别获得质粒pJF39和pJF44。用XbaI和EcoRI双酶切质粒pJF44,回收含idi(EC)基因的DNA片段,用SpeI和EcoRI双酶切质粒pJF39,将idi(EC)DNA片段与pJF39酶切产物连接,获得的质粒命名为pJF63,如图2所示。
将质粒pJF63转化重组菌E.colipJF707,获得相应的重组大肠杆菌BL21(DE3)pJF63/pJF707。
在与实施例2基本相同的条件下,对上述菌株进行培养,对发酵产物进行成分分析。结果表明,重组大肠杆菌BL21(DE3)pJF63/pJF707合成了10.7mg/L的广藿香醇,比BL21(DE3)pJF707提高了2.3倍。
实施例4
萜类合成前体供应的强化
为了进一步增强广藿香醇在重组微生物中的合成,需要对宿主的萜类前体MEP合成途径进行进一步改造。在本实施例中,首先引入异源MEP途径关键基因,构建了用于过表达枯草芽孢杆菌来源的idi基因(idi(BS))和阿维链霉菌来源的dxs2基因(dxs2(SAV))的质粒和菌株。
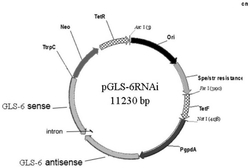
分别以枯草芽孢杆菌和阿维链霉菌的基因组DNA作为模板,用引物(idi(BS)F和idi(BS)R,dxs2(SAV)F和dxs2(SAV)R)分别PCR扩增idi(BS)、dxs2(SAV)。将纯化后的PCR产物idi(BS)和dxs2(SAV)分别用NcoI和BamHI,XbaI和HindIII双酶切后,连接到pET21d的相应酶切位点间分别获得质粒pJF554、pJF567。用XbaI和SpeI双酶切质粒pJF554,回收含idi(BS)基因的DNA片段,用SpeI单酶切质粒pJF567,将idi(BS)DNA片段与pJF567酶切产物连接。获得的重组克隆经测序鉴定,确认idi(BS)基因与dxs2(SAV)基因表达方向一致,获得的质粒命名为pJF678,如图3所示。
将质粒pJF554、pJF567和pJF678分别转化重组菌E.colipJF707,分别获得相应的重组大肠杆菌BL21(DE3)pJF554/pJF707、BL21(DE3)pJF567/pJF707和BL21(DE3)pJF678/pJF707。
在与实施例3基本相同的条件下,对上述菌株进行培养,对发酵产物进行成分分析,结果表明重组大肠杆菌BL21(DE3)pJF554/pJF707、BL21(DE3)pJF567/pJF707和BL21(DE3)pJF678/pJF707分别合成了12.32mg/L,25.63mg/L,37.25mg/L。其中共表达idi(BS)基因与dxs2(SAV)基因的菌株BL21(DE3)pJF678/pJF707最显着的改善了广藿香醇的合成,比BL21(DE3)pJF707提高了8倍。利用枯草芽孢杆菌idi(BS)和阿维链霉菌dxs2(SAV)基因共表达比过表达大肠杆菌MEP途径内源基因能够取得更明显的作用。
实施例5
萜类合成前体供应的强化:
为了进一步增强广藿香醇的微生物合成,可以在微生物细胞中同时引入最优化的MEP途径和MVA途径。在本实施例中,在过表达idi(BS)基因与dxs2(SAV)的MEP途径改造重组大肠杆菌基础上,引入了由不同其它微生物来源基因组装的高效MVA途径,构建了相关的质粒和菌株。具体地,本发明人构建了由大肠杆菌自身atoB基因,粪肠球菌mvaS(EF)和mvaE(EF)基因,金黄色葡萄球菌来源mvaK1(SAU)、mvaK2(SAU)和mvaD(SAU)基因组成的MVA途径,并将其引入了工程菌株中,进一步强化萜类前体的供应。
利用相应菌株的基因组DNA为模板,用引物atoB(EC)F和atoB(EC)R,mvaS(EF)F和mvaS(EF)R,mvaE(EF)F和mvaE(EF)R,mvaK1(SAU)F和mvaK1(SAU)R,mvaK2(SAU)F和mvaK2(SAU)R,mvaD(SAU)F和mvaD(SAU)R分别PCR扩增E.coli的atoB(EC)、粪肠球菌的mvaS(EF)和mvaE(EF)基因,金黄色葡萄球菌的mvaK1(SAU)、mvaK2(SAU)和mvaD(SAU)基因。将纯化后的PCR产物分别用NheI和HindIII,NcoI和SacI,NheI和SacI,NcoI和SacI,NcoI和SacI,NcoI和SacI连接到载体(atoB(EC)克隆到质粒pET21c(购自EMD4Biosciences)上,其它基因克隆到质粒pET28a上)的对应位点间,分别获得质粒pJF640、pJF641、pJF642、pJF645、pJF646、pJF647。然后用XbaI和XhoI双酶切质粒pJF641、pJF642,回收含mvaS(EF)、mvaE(EF)的DNA片段,并按序将其插入质粒pJF640的SpeI和XhoI位点间,获得含T7Prom-atoB(EC)-mvaS(EF)-mvaE(EF)-T7Term表达单元的质粒pJF644。
用XbaI和XhoI双酶切质粒pJF646、pJF647,回收含mvaK1(SAU)、mvaK2(SAU)的DNA片段,并按序将其插入质粒pJF643的SpeI和XhoI位点间,获得含T7Prom-mvaK1(SAU)-mvaK2(SAU)-mvaD(SAU)-T7Term的质粒pJF649。
用质粒pJF645作为模板,用引物T7PromF和mvaE(EF)2RPCR扩增T7Prom-atoB(EC)-mvaS(EF)-mvaE(EF)DNA片段。用BglII单酶切PCR产物DNA和质粒pJF649,然后将两者进行连接。获得的重组质粒经测序验证后,确认T7Prom-atoB(EC)-mvaS(EF)-mvaE(EF)与T7Prom-mvaK1(SAU)-mvaK2(SAU)-mvaD(SAU)两个表达单元的方向一致后,将重组质粒命名为pJF680,如图4所示。质粒pJF680为卡那霉素抗性,含有不同微生物来源基因的重组MVA途径。
为了能够将pJF680与含异源途径的质粒共转化E.coli,将卡那霉素抗性的质粒pJF707用XbaI和HindIII酶切,回收含pts-ispA基因的DNA片段,将其克隆到质粒pJF25(将常规的pET21c质粒的氨苄青霉素抗性基因由安普拉抗性基因替换而形成)的XbaI和HindIII位点间,获得质粒pJF708。因此该质粒携带安普拉抗性基因。
将质粒pJF680分别导入E.colipJF708菌株和强化MEP途径的E.colipJF678/pJF708菌株中,分别获得仅引入MVA途径的大肠杆菌BL21(DE3)pJF680/pJF708菌株和共同强化MEP与MVA途径的菌株BL21(DE3)pJF678/pJF680/pJF708。在与实施例3基本相同的条件下,对上述菌株进行培养,对发酵产物进行成分分析,结果表明菌株BL21(DE3)pJF680/pJF708合成了104.76mg/L广藿香醇,而菌株pJF678/pJF680/pJF708的广藿香醇的产量达到了最高的54.72mg/L(图7)。
实施例6
广藿香醇表达情况的分析
实施例2-5中部分菌株的产物合成情况汇总于下表3和图4-5中。
表3
注:5号菌株中,dxs2(SAV)+idi(BS)都是MEP途径的蛋白。
图4为工程菌株BL21(DE3)pJF707经摇瓶发酵后,发酵液由乙酸乙酯1:1萃取后,吸取有机相经GC-MS检测后的GC检测图谱,其中,广藿香醇的出峰时间为8.56min,与标准品广藿香醇的出峰时间一致。此外,8.56min峰的质谱图也与广藿香醇的标准品质谱图一致(图5)。这表明在重组大肠杆菌中成功地合成了广藿香醇。
经过5天的摇瓶培养后,菌株BL21(DE3)pJF707能够合成约4.65mgl-1的广藿香醇,如图7。同时合成的产物中还含有其它的多种萜类副产物。前人的研究表明过表达某些植物源或者其它微生物来源的MEP途径的dxs,idi基因比过表达E.coli内源的基因能够取得更显着地效果。本研究选取了S.avermitilis来源的dxs2(SAV)基因和B.subtilis的二型idi(BS)基因进行过表达。单独过表达dxs2(SAV)和idi(BS)的工程菌株BL21(DE3)pJF567/pJF707和pJF554/pJF707能够合成12.32mg/L和25.63mg/L的广藿香醇。经5天发酵后,该产量均明显超过了BL21(DE3)pJF63/pJF707。而共表达dxs2(SAV)和idi(BS)的工程菌株BL21(DE3)pJF678/pJF707能够合成37.25mgl-1的广藿香醇,是采用E.coli内源模块的工程菌株BL21(DE3)pJF63/pJF707的3.48倍。
本发明人组装了由E.coli来源的atoB基因,粪肠球菌的mvaS和mvaE基因,金黄色葡萄球菌mvaK1、mvaK2和mvaD基因组成的外源MVA途径,并将其与外源MEP途径模块一起在工程菌株中进行共表达,来进一步广藿香醇的产量。工程菌株BL21(DE3)pJF680/pJF708能够合成104.76mg/L的广藿香醇,而同时进行MEP途径强化和引入MVA途径的菌株BL21(DE3)pJF680/pJF678/pJF708的产量仅为54.72mg/L。研究结果表明,E.coli工程菌株能够高效地合成广藿香醇,并为进一步提高广藿香醇的产量打下了基础。
讨论
采用代谢工程技术,在操作简单、易培养的微生物细胞(如大肠杆菌Escherichiacoli、酿酒酵母Saccharomycescerevisiae)中重构不同植物源萜类化合物的下游合成途径,结合对工程菌株前体合成的遗传改造是高效地生产这些萜类化合物的重要方法。
2-C-甲基-D-赤藻糖醇-4-磷酸(2-C-methyl-D-erythritol-4-phosphate,MEP)途径和甲羟戊酸(Mevalonate,MVA)途径是自然界中存在的两条用于萜类化合物合成的途径。前者主要存在于原核细菌(如大肠杆菌Escherichiacoli),植物质体等,后者主要存在于真核细胞中。
本发明人通过对工程菌株的MEP途径和MVA途径进行代谢工程改造。实验结果表明,这样可进一步显着地提高广霍香醇化合物的产量。
在本发明提及的所有文献都在本申请中引用作为参考,就如同每一篇文献被单独引用作为参考那样。此外应理解,在阅读了本发明的上述讲授内容之后,本领域技术人员可以对本发明作各种改动或修改,这些等价形式同样落于本申请所附权利要求书所限定的范围。
异源合成广霍香醇的方法和微生物专利购买费用说明
![]()
Q:办理专利转让的流程及所需资料
A:专利权人变更需要办理著录项目变更手续,有代理机构的,变更手续应当由代理机构办理。
1:专利变更应当使用专利局统一制作的“著录项目变更申报书”提出。
2:按规定缴纳著录项目变更手续费。
3:同时提交相关证明文件原件。
4:专利权转移的,变更后的专利权人委托新专利代理机构的,应当提交变更后的全体专利申请人签字或者盖章的委托书。
Q:专利著录项目变更费用如何缴交
A:(1)直接到国家知识产权局受理大厅收费窗口缴纳,(2)通过代办处缴纳,(3)通过邮局或者银行汇款,更多缴纳方式
Q:专利转让变更,多久能出结果
A:著录项目变更请求书递交后,一般1-2个月左右就会收到通知,国家知识产权局会下达《转让手续合格通知书》。













动态评分
0.0