

IPC分类号 : C12N9/90,C12N15/61,C12P15/00,C12P17/18,C12P17/04



专利摘要
一种新的雷公藤柯巴基焦磷酸合酶TwCPS4及其编码基因,TwCPS4在诱导初期雷公藤二萜化合物的合成过程中起重要作用。催化实验表明TwCPS4与丹参SmKSL1联合使用,能够以GGPP为底物合成松香烷型二萜前体,证实了TwCPS4参与雷公藤二萜类次生合成途径。TwCPS4与丹参SmKSL1组合合成松香烷型二萜前体的效率高于SmCPS1与SmKSL1的组合。
权利要求
1.雷公藤柯巴基焦磷酸合酶TwCPS4,其氨基酸序列由SEQ ID NO:1所示。
2.编码权利要求1所述雷公藤柯巴基焦磷酸合酶TwCPS4的基因,其核苷酸序列由SEQ ID NO:2所示。
3.雷公藤柯巴基焦磷酸合酶TwCPS4或其基因在合成二萜类化合物中的用途。
4.如权利要求3所述用途,其特征在于雷公藤柯巴基焦磷酸合酶TwCPS4或其基因,与丹参贝壳烯合酶SmKSL1或其基因联合使用合成松香烷型二萜类化合物。
5.如权利要求3或4所述用途,其特征在于所述二萜类化合物包括雷公藤甲素或其前体、丹参酮或其前体等,优选的,所述二萜类化合物是次丹参酮二烯。
6.如权利要求1雷公藤柯巴基焦磷酸合酶TwCPS4或其基因在合成松香烷型二萜类化合物中的用途,其特征在于先使用雷公藤柯巴基焦磷酸合酶TwCPS4以GGPP为底物催化产生CPP,再使用丹参贝壳烯合酶SmKSL1催化CPP合成松香烷型二萜类化合物前体。
7.一种合成松香烷型二萜类化合物前体的方法,其包括:
(1)将雷公藤柯巴基焦磷酸合酶TwCPS4的编码基因表达盒插入原核表达载体中,制备表达外源雷公藤柯巴基焦磷酸合酶TwCPS4的重组大肠杆菌,超声破碎菌体,取含有重组TwCPS4蛋白的上清液;
(2)将丹参贝壳烯合酶SmKSL1的编码基因表达盒插入原核表达载体中,制备表达外源丹参贝壳烯合酶SmKSL1的重组大肠杆菌,超声破碎菌体,取含有重组SmKSL1蛋白的上清液;
(3)以GGPP为底物,以步骤(1)中获得的含重组TwCPS4蛋白的上清液为催化剂,催化GGPP产生CPP;
(4)以步骤(3)获得的CPP为底物,以步骤(2)获得的含重组SmKSL1蛋白的上清液为催化剂,催化CPP产生松香烷型二萜类化合物前体。
8.如权利要求7所述合成松香烷型二萜类化合物前体的方法,步骤(1)所述的原核表达载体为pMAL-c2X,所述TwCPS4基因插入BamH I和Sal I之间。
9.如权利要求7所述合成松香烷型二萜类化合物前体的方法,在步骤(3)的催化反应完成后还包括正己烷萃取、脱磷的步骤。
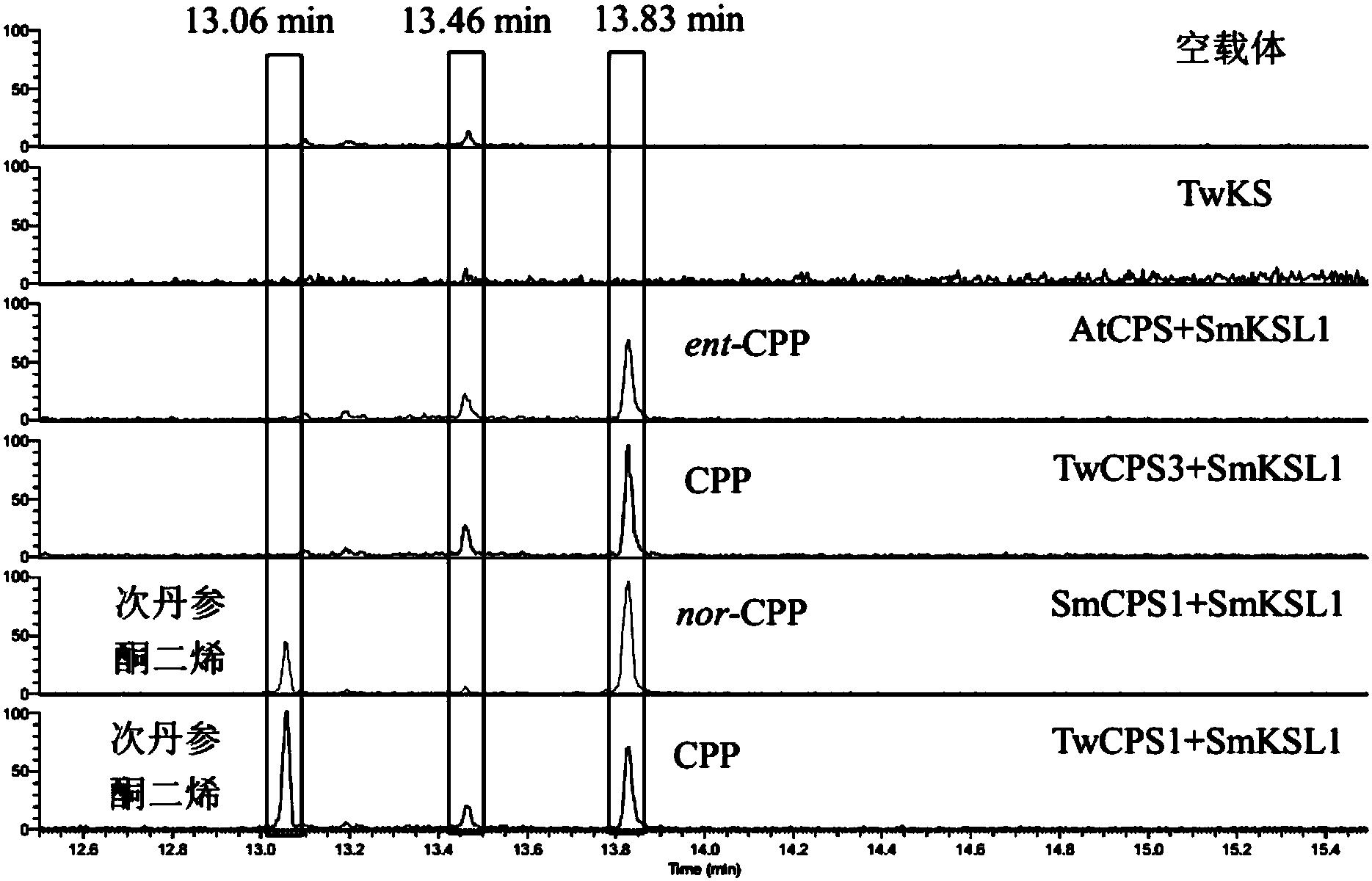
10.如权利要求7所述合成松香烷型二萜类化合物前体的方法,在步骤(4)完成后还包括对反应产物进行GC-MS分析的步骤,其中在第13.06分出现的峰为次丹参酮二烯。
说明书
技术领域
本发明属于生物制药领域,具体涉及通过基因工程方法获得二萜类化合物合成的关键酶,特别涉及雷公藤二萜合酶及其在松香烷型二萜化合物制备中的用途。
背景技术
药用植物雷公藤(Tripterygium wilfordii.Hook.f.)是一味中草药,
被广泛用于类风湿性关节炎和炎症的治疗(Raphaela G M,Mildred W,Roy F,et al.Comparison of Tripterygium wilfordii Hook F Versus Sulfasalazine in the Treatment of Rheumatoid Arthritis:A Randomized Trial[J].Annals of Internal Medicine,2009,151(4):229-240.Tao X L,Lipsky P E.The Chinese anti-inflammatory and immunosuppressive herbal remedy Tripterygium wilfordii Hook F.[J].Rheumatic Disease Clinics of North America,2000,26(1):29-50.)。萜类成分为雷公藤的主要活性成分,包括雷公藤甲素(triptolide)、雷酚内酯(triptophenolide)和雷公藤红素(celastrol)等。从中药中的活性成分开发新药是一种很有潜力的方式,然而由于植物的生长缓慢,再加上这些有效成分在植物体中的含量不多,因而大大限制了它的发展。通过探寻和阐释萜类成分在雷公藤中的生物合成途径及其调控机制,有助于为药材品质的形成提供理论基础,同时为利用生物技术提高目标成分含量或直接生产有效成分或中间体带来广阔的应用空间。
二萜类化合物通常以GGPP为底物,在二萜合酶复合体的催化作用下形成,即IPP通过一系列的反应形成GGPP后,二萜合酶复合体以GGPP为起始底物,经过最初的离子异构化后环化形成各种阳离子中间产物,最终通过去质子或捕获亲核物质进一步环化形成一系列的二萜骨架中间体;并通过细胞色素P450结构修饰,主要包括羟基化、甲基化、异构化、脱甲基化、加成和还原等,最终生成二萜类化合物。
雷公藤中二萜类化合物结构复杂、种类繁多,合成代谢途径尚不完全清楚。目前基于基因组序列分析和表达序列标签分析,对雷公藤二萜合酶复合体中的部分酶活性蛋白进行了初步预测,但远远不能满足雷公藤二萜化合物生物合成的需求,不能解释雷公藤二萜化合物种类的多样性,更不能确定预测的酶活性蛋白具体导致了何种二萜类产物,因此迫切需要挖掘雷公藤二萜合酶复合体中新的酶活蛋白,并确定其具体的催化功能。
雷公藤甲素具有抗肿瘤、抗炎、抗排异、抗生育等药理作用,雷公藤甲素与丹参酮类化合物同属松香烷型二萜。酶催化制备丹参酮的方法已有报道,但酶催化制备雷公藤甲素尚未获得成功。针对雷公藤甲素、丹参酮共同的松香烷型二萜化合物前体进行研究能够为最终酶催化法合成雷公藤甲素打下基础。
发明内容
发明人团队在对雷公藤二萜合酶复合体中的酶活蛋白进行研究过程中,意外的发现了一种与TwCPS1序列相似的柯巴基焦磷酸合酶。最初认为其是TwCPS1的天然突变等效物,随后进一步的研究表明其与TwCPS1同时稳定存在于同一株雷公藤悬浮细胞中,并且二者在经茉莉酸甲酯诱导时的过表达情况也不同。新发现的柯巴基焦磷酸合酶对诱导的反应更快,负责诱导初期松香烷型二萜化合物的合成和累积,而TwCPS1对诱导的反应滞后,负责诱导中后期松香烷型二萜化合物的合成和累积,因此将这种新的柯巴基焦磷酸合酶命名为TwCPS4。
将克隆得到的雷公藤柯巴基焦磷酸合酶基因Twcps4和丹参中的Smksl1基因分别构建表达载体,成功生成了次丹参酮二烯。进一步证实了TwCPS4在合成雷公藤甲素等松香烷型二萜合成中的作用,且提供了一种合成松香烷型二萜前体的方法,为雷公藤甲素的生物合成奠定基础。
次丹参酮二烯是一种重要的松香烷型二萜化合物前体,由于其发现于丹参,因此通常将丹参中的SmCPS1与SmKS1用于制备次丹参酮二烯。除了丹参以外,其他的植物中尽管也存在二萜合酶KS、CPS,理论上也可能产生二萜类化合物,但通常不能催化合成次丹参酮二烯或合成量低于丹参自身SmCPS1与SmKSL1的组合。本发明提供的雷公藤柯巴基焦磷酸合酶TwCPS4替代丹参SmCPS1,与SmKSL1组合使用能够成功合成次丹参酮二烯,且产量更高。即:本发明所述雷公藤二萜合酶TwCPS4能够用于生产松香烷型二萜化合物前体,并且代替丹参二萜合酶SmCPS1能获得更高的产量,在催化合成松香烷型二萜化合物前体中具有广阔的应用前景。
本发明提供了一种合成雷公藤松香烷型二萜类化合物前体合成的方法:具体将Twcps4基因的eDNA克隆到原核表达载体pMAL-c2X,构建带有Twcps4基因的重组表达载体;转入E.coli表达宿主菌,通过诱导,表达产物经初步纯化后,加入到以GGPP为底物的体外酶促反应体系中,正己烷萃取反应产物,采用GC-MS可检测到产物。研究结果表明,本发明与雷公藤二萜类化合物合成有关的基因具有二萜合酶基因的DXDD特征结构域,酶促反应分析发现该基因催化雷公藤二萜类化合物的早期合成步骤,对于培育高品质的药用植物品种特别是培育具有高雷公藤甲素含量的雷公藤品种具有重要的理论及实际意义。
本发明提供一种雷公藤柯巴基焦磷酸合酶TwCPS4,其氨基酸序列由SEQ ID NO:1所示。
本发明还提供编码所述雷公藤柯巴基焦磷酸合酶TwCPS4的基因,其核酸序列由SEQ ID NO:2所示。
本发明还提供雷公藤柯巴基焦磷酸合酶TwCPS4或其基因在合成二萜类化合物中的用途。
本发明所述雷公藤柯巴基焦磷酸合酶TwCPS4或其基因在合成二萜类化合物中的用途,其中雷公藤柯巴基焦磷酸合酶TwCPS4或其基因,与丹参贝壳烯合酶SmKSL1或其基因联合使用合成松香烷型二萜类化合物。
本发明所述雷公藤柯巴基焦磷酸合酶TwCPS4或其基因在合成二萜类化合物中的用途,其中所述二萜类化合物包括雷公藤甲素或其前体、丹参酮或其前体等,优选的,所述二萜类化合物是次丹参酮二烯。
本发明所述雷公藤柯巴基焦磷酸合酶TwCPS4或其基因在合成二萜类化合物中的用途,其中先使用雷公藤柯巴基焦磷酸合酶TwCPS4以GGPP为底物催化产生CPP,再使用丹参贝壳烯合酶SmKSL1催化所产生的CPP合成松香烷型二萜类化合物前体。
本发明还提供一种合成松香烷型二萜类化合物前体的方法,其包括:(1)将雷公藤柯巴基焦磷酸合酶TwCPS4的编码基因表达盒插入原核表达载体中,制备表达外源雷公藤柯巴基焦磷酸合酶TwCPS4的重组大肠杆菌,超声破碎菌体,取含有重组TwCPS4蛋白的上清液;
(2)将丹参贝壳烯合酶SmKSL1的编码基因表达盒插入原核表达载体中,制备表达外源丹参贝壳烯合酶SmKSL1的重组大肠杆菌,超声破碎菌体,取含有重组SmKSL1蛋白的上清液;
(3)以GGPP为底物,以步骤(1)中获得的含重组TwCPS4蛋白的上清液为催化剂,催化GGPP产生CPP;
(4)以步骤(3)获得的CPP为底物,以步骤(2)获得的含重组SmKSL蛋白的上清液为催化剂,催化CPP产生松香烷型二萜类化合物前体。
本发明所述合成松香烷型二萜类化合物前体的方法,步骤(1)所述的原核表达载体为pMAL-c2X,所述Twcps4基因插入BamH I和Sal I之间。
本发明所述合成松香烷型二萜类化合物前体的方法,在步骤(3)的催化反应完成后还包括正己烷萃取、脱磷的步骤。
本发明所述合成松香烷型二萜类化合物前体的方法,在步骤(4)完成后还包括对反应产物进行GC-MS分析的步骤,其中在第13.06分出现的峰为次丹参酮二烯。
附图说明
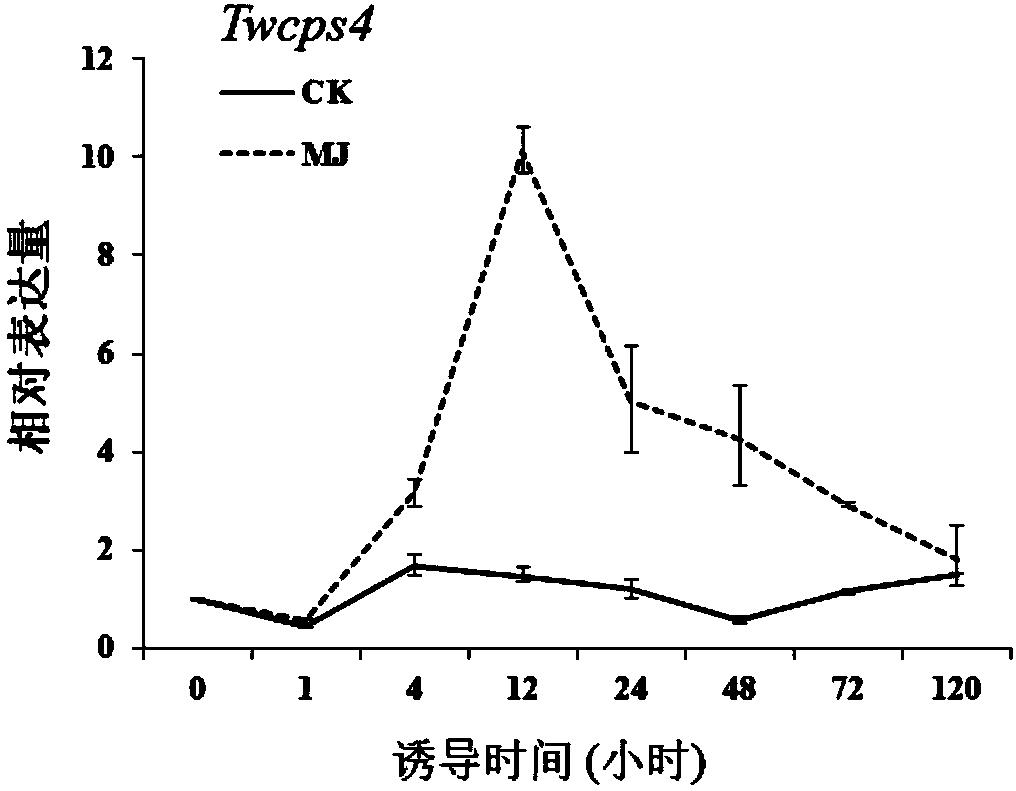
图1茉莉酸甲酯诱导下TwCPS4不同时间的相对表达量;
图2宿主菌E.coli TransB(DE3)诱导表达产物SDS-PAGE蛋白电泳分析。M:蛋白分子量标准,条带由上往下分别为200、116、97.2、66.4KDa;箭头表示目的重组蛋白约130-140KDa;
图3 CPS催化底物GGPP形成产物提取离子色谱图。第13.83分的峰为CPP;
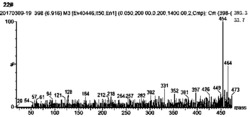
图4 CPS催化底物GGPP形成产物质谱图,分子离子峰275(m/z)为CPP,图4A为TwCPS4,图4B为SmCPS1,图4C为AtCPS;
图5(CPS+KS)催化GGPP形成产物提取离子色谱图。第13.06分的峰为次丹参酮二烯;
图6(CPS+KS)催化GGPP形成产物质谱图,分子离子峰272(m/z)为次丹参酮二烯,图A为SmCPS1+SmKSL1,图6B为TwCPS4+SmKSL1。
具体实施方式
以下结合具体实施例,进一步阐述本发明。这些实施例仅用于说明本发明而不用于限制本发明的范围。下列实施例中未注明具体条件的实验方法,通常按照常规条件或按照制造厂商所建议的条件。除非另行定义,文中所使用的所有专业与科学用语与本领域熟练人员所熟悉的意义相同。此外,任何与所记载内容相似或均等的方法及材料皆可应用于本发明方法中。文中所述的较佳实施方法与材料仅作示范之用。
下述实施例中所使用的实验方法如无特殊说明,均为常规方法。所用的材料、试剂等,如无特殊说明,均可从商业途径得到。
雷公藤(Tripterygium wilfordii Hook.F.)悬浮细胞在文献“雷公藤4-(5’-二磷酸胞苷)-2-C-甲基-D-赤藓醇激酶基因的全长克隆与表达分析.中国中药杂志,2015,40(21):4165-4170”中公开过,公众可从首都医科大学分子生药与中药资源实验室获得。
SMARTerTM RACE cDNA Amplification Kit,PrimeSTAR GXL DNA Polymerase购自Takara公司;pEASY-Blunt Simple Cloning Kit购自北京全式金生物技术有限公司,GGPP购自Sigma公司,产品目录号为G6025。
实施例一、雷公藤TwCPS4诱导表达分析
根据雷公藤管家基因β-actin和雷公藤Twcps4基因的核苷酸序列设计引物。待目的基因引物及管家基因引物检测合格后,在ABI 7500型实时荧光定量PCR仪上进行实时荧光定量PCR,配备体系如下:
PCR反应条件为:
所述实时荧光定量PCR引物序列如下:
反应结束后分析荧光值变化曲线和融解曲线。每个反应重复3次,采用2-ΔΔCT法分析结果。
根据实时荧光定量PCR实验检测结果数据,采用2-ΔΔCT方法进行相对定量表达分析。实验发现MJ能明显诱导雷公藤悬浮细胞中Twcps1、Twcps2、Twcps3、Twcps4、Twks、Twges1、Twges2基因mRNA的表达。具体表现为MJ处理后的Twcps2、Twcps3、Twcps4基因的诱导表达水平在12h内均迅速达到最大值;Twcps1、Twks基因基因的诱导表达水平在24h内均迅速达到最大值;Twges1、Twges2基因的诱导表达水平在120h内均迅速达到最大值;MJ处理组Twcps1基因的表达量是同期对照组的23.03倍;MJ处理组Twcps2基因的表达量是同期对照组的30.08倍;MJ处理组Twcps3基因的表达量是同期对照组的6.77倍;MJ处理组Twcps4基因的表达量是同期对照组的6.89倍(图1);MJ处理组Twks基因的表达量是同期对照组的35.91倍。
实施例二、雷公藤Twcps4原核表达载体构建
以含有雷公藤Twcps4基因全长cDNA的载体pMD-19-T-CPS4质粒为模板,用含酶切位点引物,进行PCR扩增基因编码区(引物序列见表3-1)。DNA聚合酶采用高保真DNA聚合酶(PrimeSTAR HS DNA Polymerase)。PCR参数为98℃ 3min,1循环;98℃ 10s,60℃ 10s,72℃ 2min30s,30循环;72℃ 7min;4℃维持。扩增产物经Gene JET Gel Extraction Kit胶回收(方法如下)。
Gene JET Gel Extraction Kit胶回收步骤:
(1)取PCR产物与6×loading buffer预混合,在1.5%琼脂糖凝胶上以低电压(约5Vcm-1)电泳30-60min;
(2)用解剖刀或剃刀片切割含有DNA片段的凝胶,尽可能的靠近DNA片段切割,以减小凝胶的含量,将胶片放在事先称重的1.5mL离心管并称重。记录胶片的重量。注意避免长时间暴露在紫外灯下,损害DNA而影响后续实验;
(3)加1∶1量的Binding Buffer到胶片中(量以重量计,如每100毫克琼脂糖凝胶加100微升的Binding Buffer);
(4)在50-60℃的条件下温育凝胶混合物10min,期间颠倒混匀2-3次,促进胶融化,保证胶全部溶解,在上柱前将凝胶混合物快速涡旋混匀一次;
(5)转移最多800μL凝胶溶解液到基因回收纯化柱,13000g离心1min,弃去流出液,然后将柱放回相同的收集管;
(6)加入700μLWash Buffer(已用乙醇稀释)到Gene JET纯化柱。13000g离心1min,弃去流出液,然后将柱放回相同的收集管;
(7)离心空GeneJET纯化柱13000g离心1min,彻底去除残留的Wash Buffer;
(8)将Gene JET纯化柱转移到一个干净的1.5mL离心管,加30-50μL ddH20(可60℃预热)于纯化柱膜,13000g离心1min;
(9)丢掉Gene JET纯化柱并储存纯化的DNA在-20℃。
经纯化后的PCR产物用限制性内切酶进行双酶切,采用NEB公司T4DNA连接酶定向连入经相同双酶切的表达载体pMAL-c2X中(方法如下),连接产物经转化、蓝白菌落筛选及阳性克隆的初步筛选,并送样测序鉴定,得到经测序核苷酸序列无突变重组质粒pMALTPS,并将经的重组质粒pMALTPS转化至大肠杆菌E.coli TransB(DE3)表达感受态细胞。与此同时,将拟南芥中功能明确的AtCPS(催化GGPP形成ent-CPP)、AtKS(催化ent-CPP形成对映贝壳杉烯)及丹参中功能明确的SmCPS1(催化GGPP形成nor-CPP)、SmKSL1(催化nor-CPP形成次丹参酮二烯)构建到pMAL-c2X原核表达载体上(SmCPS1、SmKSL1利用实验室保存的重组质粒pET32SmCPS1及pET32SmKSL1),将雷公藤TwCPS1、TwCPS2、TwCPS3、TwKS与其他植物来源的CPS、KS进行对比。
采用TaKaRa QuickCut酶进行双酶切反应:
双酶切反应体系
双酶切反应(50μL体系)
内切酶1酶切完成后,加入1μL内切酶2继续酶切。注意:最适酶切温度较低的内切酶应先进行酶切,若最适酶切温度相同,可同时加入同时酶切。
采用NEB T4 DNA快速连接试剂盒进行DNA片段连接反应:
连接反应体系
连接反应(20μL体系)
*DNA与载体摩尔比约为3∶1-10∶1
25℃连接5min(可适当延长连接时间)。
构建原核表达载体引物序列
实施例三、重组蛋白诱导表达
诱导表达
100mmol·L-1IPTG:称取238.3mg的IPTG用10mL的ddH2O溶解,过滤分装-20℃保存;LB培养基:Trytone 1.0%,Yeast Extract 0.5%,NaCl1.0%,Agar1.5%,pH 7.0。
操作步骤:
(1)将酶切鉴定正确且经测序验证的含目的基因的重组质粒,取1μL转化至50μl TransB(DE3)感受态细胞中,涂板LB+Amp(氨苄青霉素钠)固体平板,37℃倒置培养12-16h;
(2)挑取单克隆菌落经酶切鉴定后转到含100μg·mL-1Amp的2mL LB液体培养基中,37℃振荡培养至OD600到0.6-1.0;
(3)取1mL菌液5000g4℃离心5min收集菌体,用新鲜LB+Amp液体培养基混悬菌体,转接到100mL LB+Amp液体培养基中;
(4)37℃培养至宿主菌密度(OD600)达到0.6-1.0时,加入适量IPTG诱导剂(终浓度约0.4mM),在低温下(16℃)诱导培养8h;
(5)4℃ 3000g离心20min收获菌体,预冷的5mL HEPES缓冲液(50mM HEPES,100mM KCl,7.5mM MgCl2,5mM DTT,1mM PMSF,5%甘油,pH 7.2)重悬;
(6)置冰浴中超声破菌(30%功率,超声5s,间隔5s,持续3min);大肠杆菌破碎液在4℃,15000g离心30min,取上清液;
(7)上清液经蛋白超滤管浓缩(4℃ 5000g 40min)至1.5mL。
取上清进行十二烷基硫酸钠-聚丙烯酰胺(SDS-PAGE)凝胶电泳检测。
实施例四、十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(SDS-PAGE)(图2)
试剂配制
30%丙烯酰胺贮存液(神经毒性,操作时配戴口罩及手套):在通风橱中,称取丙烯酰胺29.2g,甲叉双丙烯酰胺0.8g,加ddH2O溶解后,定容到100mL。针头滤器过滤后置棕色瓶中,4℃保存;
pH 8.8Tris-HCl分离胶缓冲液:配制1.5M Tris-HCl,并调节pH至8.8,4℃保存;
pH 6.8Tris-HCl浓缩胶缓冲液:配制1M Tris-HCl,并调节pH至6.8,4℃保存;
10%SDS:称取SDS 1.0g,蒸馏水10mL溶解,4℃保存;
10%过硫酸铵(APS):取APS 1.0g,蒸馏水10mL溶解,4℃保存;
TEMED(四乙基乙二胺)原液;
5×样品缓冲液(10mL):0.6mL 1mol·L-1的Tris-HCl(pH 6.8),5mL 50%甘油,2mL 10%的SDS,0.5mL巯基乙醇,1mL 1%溴酚蓝,0.9mL蒸馏水。可在4℃保存数周,或在-20℃保存数月;
10×电泳缓冲液:称取Tris 30.38g,甘氨酸144g,SDS 10.8g,加蒸馏水约900mL,调pH 8.3后,用蒸馏水定容至1000mL,置4℃保存,临用前稀释10倍。
实施例五、样品制备
将蛋白质样品与5×样品缓冲液在一个Eppendorf管中混合,放入100℃加热5-10min,常温12000g离心3min,取上清点样。
实施例六、电泳
操作步骤
(1)将玻璃板、样品梳用洗涤剂洗净,用ddH2O冲洗数次,晾干;
(2)安装玻璃板;
(3)按如下体积配制10%分离胶15mL(配制两块胶),混匀;
(4)向玻璃板间灌制分离胶,立即加入1mL ddH20压平胶面,大约20min后胶即可聚合;
(5)按如下体积配制5%浓缩胶5mL(配制两块胶),混匀;
(6)将上层ddH2O倾去,滤纸吸干,灌制浓缩胶,插入样品梳;
(7)装好电泳系统,加入电泳缓冲液,上样5-10μL;
(8)恒流40mA,溴酚蓝跑出分离胶30min后,停止电泳;
(9)卸下胶板,剥离胶,染色,照胶。
实施例七、考马斯亮蓝染色
试剂配制
考马斯亮蓝R250染色液:0.25%考马斯亮蓝R250(W/V),45%甲醇(V/V),10%冰乙酸;
脱色液(1000mL):100m冰乙酸,250mL乙醇,蒸馏水补足至1000mL。
染色步骤
室温染色45min-60min(或微波炉快染,高火20s,两次);用蒸馏水清洗3-5遍;加入脱色液,置于100rpm摇床上脱色,每20min更换一次脱色液至胶透明,脱色完成后,用UMAX Powerlook 2100XL扫描仪进行照胶。
实施例八、酶促反应
以GGPP为底物的酶促反应,取实施例三、步骤(7)获得的浓缩重组CPS蛋白上清进行酶促反应。并以pMAL-c2X空载体、pMAL-c2X-AtCPS、pET32a(+)-SmCPS1转化大肠杆菌TransB(DE3)表达感受态细胞,所破碎浓缩得到的重组蛋白上清进行酶促反应作为对照。
酶促反应体系如下:
浓缩重组蛋白上清182μL
底物GGPP(200μM) 18μL
反应产物充分混合(枪头吹打),在室温(25℃)、黑暗的环境下反应2h;反应结束后用正己烷抽提3次,每次加入0.5mL,弃有机相,留水相(留一层正己烷覆盖水相,防止产物氧化);然后用N2将水相中的正己烷彻底吹干,以免影响下一步的脱磷反应。
脱磷反应体系如下:
反应产物充分混合(枪头吹打),在37℃下反应4h;脱磷后的产物再用正己烷抽提3次,每次加入0.5mL,将抽提所得有机相汇合在一起;用N2将提取液吹干,并加入60μL正己烷溶解,用于GC-MS分析(图3、图4)。
以CPP为底物的酶促反应
对TwKS进行功能分析的酶促反应是以经CPS酶催化GGPP所产生的产物(CPP)作为反应底物,分析TwKS酶催化所得产物的化学结构,从而鉴定TwKS功能。具体实验过程如下:
(1)配制如下酶促反应体系,在室温(25℃)黑暗条件下反应2小时,使GGPP充分转化成CPP;
浓缩重组KS蛋白上清182μL
底物GGPP(200μM) 18μL
(2)向上述反应混合液中加入等体积的TwKS酶,并补充MgCl2至10mM,在室温(25℃)黑暗条件下,反应过夜(12-16h);
(3)反应结束后,体系用正己烷抽提3次(每次0.5mL),所得正己烷相用N2吹干,然后加入60μL正己烷溶解进行GC-MS分析(图5、6)。实施例九、反应产物GC-MS检测
GC-MS分析条件为:取1μL的进样,无分流的模式下,50℃保持2min,20℃·min-1升至300℃,保持20min;进样口温度250℃,离子源温度250℃,电子能量70ev,对样品进行20-650m/z范围扫描。GC-MS仪器为Thermo Scientific公司Thermo TRACE 1310/TSQ 8000gas chromatograph,色谱柱为DB-5ms(30m×0.25mm)。
上述说明并非对本发明的限制,本发明也并不限于上述举例。本技术领域的普通技术人员在本发明的实质范围内,作出的变化、改型、添加或替换,也应属于本发明的保护范围,本发明的保护范围以权利要求书为准。
SEQUENCE LISTING
<110> 首都医科大学
<120> 雷公藤焦磷酸合酶TwCPS4及其制备松香烷型二萜化合物中的应用
<130> 无
<160> 4
<170> PatentIn version 3.5
<210> 1
<211> 806
<212> PRT
<213> 雷公藤
<400> 1
Met His Ser Leu Leu Met Lys Lys Val Ile Met Tyr Ser Ser Gln Thr
1 5 1015
Thr His Val Phe Pro Ser Pro Leu His Cys Thr Ile Pro Lys Ser Ser
202530
Ser Phe Phe Leu Asp Ala Pro Val Ala Arg Leu His Cys Leu Ser Gly
354045
His Gly Ala Lys Lys Lys Arg Leu His Phe Asp Ile Gln Gln Gly Arg
505560
Asn Ala Ile Ser Lys Thr His Thr Pro Asp Leu Tyr Ala Lys Gln Glu
65707580
Tyr Ser Val Pro Glu Ile Val Lys Gly Asp Asp Lys Glu Glu Glu Val
859095
Val Lys Ile Lys Glu His Val Asp Ile Ile Lys Ser Met Leu Ser Ser
100 105 110
Met Glu Asp Gly Glu Ile Ser Ile Ser Ala Tyr Asp Thr Ala Trp Val
115 120 125
Ala Leu Ile Gln Asp Ile His Asn Asn Gly Ala Pro Gln Phe Pro Ser
130 135 140
Ser Leu Leu Trp Ile Ala Glu Asn Gln Leu Pro Asp Gly Ser Trp Gly
145 150 155 160
Asp Ser Arg Val Phe Leu Ala Phe Asp Arg Ile Ile Asn Thr Leu Ala
165 170 175
Cys Val Val Ala Leu Lys Ser Trp Asn Val His Pro Asp Lys Cys Glu
180 185 190
Arg Gly Ile Ser Phe Leu Lys Glu Asn Ile Ser Met Leu Glu Lys Asp
195 200 205
Asp Ser Glu His Met Leu Val Gly Phe Glu Phe Gly Phe Pro Val Leu
210 215 220
Leu Asp Met Ala Arg Arg Leu Gly Ile Asp Val Pro Asp Asp Ser Pro
225 230 235 240
Phe Leu Gln Glu Ile Tyr Val Gln Arg Asp Leu Lys Leu Lys Arg Ile
245 250 255
Pro Lys Asp Ile Leu His Asn Val Pro Thr Thr Leu Leu His Ser Leu
260 265 270
Glu Ala Ile Pro Asp Leu Asp Trp Thr Lys Leu Leu Lys Leu Gln Cys
275 280 285
Gln Asp Gly Ser Leu Leu Phe Ser Pro Ser Ser Thr Ala Met Ala Phe
290 295 300
Ile Asn Thr Lys Asp Glu Asn Cys Leu Arg Tyr Leu Asn Tyr Val Val
305 310 315 320
Gln Arg Phe Asn Gly Gly Ala Pro Thr Val Tyr Pro Tyr Asp Leu Phe
325 330 335
Glu His Asn Trp Ala Val Asp Arg Leu Gln Arg Leu Gly Ile Ser Arg
340 345 350
Phe Phe Gln Pro Glu Ile Arg Glu Cys Met Ser Tyr Val Tyr Arg Tyr
355 360 365
Trp Thr Lys Asp Gly Ile Phe Cys Thr Arg Asn Ser Arg Val His Asp
370 375 380
Val Asp Asp Thr Ala Met Gly Phe Arg Leu Leu Arg Leu His Gly Tyr
385 390 395 400
Glu Val His Pro Asp Ala Phe Arg Gln Phe Lys Lys Gly Cys Glu Phe
405 410 415
Ile Cys Tyr Glu Gly Gln Ser His Pro Thr Val Thr Val Met Tyr Asn
420 425 430
Leu Tyr Arg Ala Ser Gln Leu Met Phe Pro Glu Glu Lys Ile Leu Asp
435 440 445
Glu Ala Lys Gln Phe Thr Glu Lys Phe Leu Gly Glu Lys Arg Ser Ala
450 455 460
Asn Lys Leu Leu Asp Lys Trp Ile Ile Thr Lys Asp Leu Pro Gly Glu
465 470 475 480
Val Gly Phe Ala Leu Asp Val Pro Trp Tyr Ala Ser Leu Pro Arg Val
485 490 495
Glu Ala Arg Phe Phe Ile Gln His Tyr Gly Gly Glu Asp Asp Val Trp
500 505 510
Leu Asp Lys Ala Leu Tyr Arg Met Pro Tyr Val Asn Asn Asn Val Tyr
515 520 525
Leu Glu Leu Ala Lys Leu Asp Tyr Asn Tyr Cys Gln Ala Leu His Arg
530 535 540
Thr Glu Trp Gly Arg Ile Gln Lys Trp Tyr Glu Glu Ser Lys Pro Arg
545 550 555 560
Asp Phe Gly Ile Ser Arg Glu Cys Leu Leu Arg Ala Tyr Phe Met Ala
565 570 575
Ala Ala Ser Ile Phe Glu Pro Glu Arg Ser Met Glu Arg Leu Ala Trp
580 585 590
Ala Lys Thr Ala Ile Leu Leu Glu Ile Ile Val Ser Tyr Phe Asn Glu
595 600 605
Val Gly Asn Ser Thr Glu Gln Arg Ile Ala Phe Thr Thr Glu Phe Ser
610 615 620
Ile Arg Ala Ser Pro Met Gly Gly Tyr Ile Asn Gly Arg Lys Leu Asp
625 630 635 640
Lys Ile Gly Thr Thr Gln Glu Leu Ile Gln Met Leu Leu Ala Thr Ile
645 650 655
Asp Gln Phe Ser Gln Asp Ala Phe Ala Ala Tyr Gly His Asp Ile Thr
660 665 670
Arg His Leu His Asn Pro Trp Lys Met Trp Leu Leu Lys Trp Gln Glu
675 680 685
Glu Gly Asp Arg Trp Leu Gly Glu Ala Glu Leu Leu Ile Gln Thr Ile
690 695 700
Asn Leu Met Ala Asp His Lys Ile Ala Glu Lys Leu Phe Met Gly His
705 710 715 720
Thr Asn Tyr Glu Gln Leu Phe Ser Leu Thr Asn Lys Val Cys Tyr Ser
725 730 735
Leu Gly His His Glu Leu Gln Asn Asn Arg Glu Leu Glu His Asp Met
740 745 750
Gln Arg Leu Val Gln Leu Val Leu Thr Asn Ser Ser Asp Gly Ile Asp
755 760 765
Ser Asp Ile Lys Lys Thr Phe Leu Ala Val Ala Lys Arg Phe Tyr Tyr
770 775 780
Thr Ala Phe Val Asp Pro Glu Thr Val Asn Val His Ile Ala Lys Val
785 790 795 800
Leu Phe Glu Arg Val Asp
805
<210> 2
<211> 2421
<212> DNA
<213> 雷公藤
<400> 2
atgcatagtt tgttaatgaa aaaggtgatc atgtactcct ctcaaacaac ccacgtcttc 60
ccttcccctc tccactgcac cattccaaaa tcttcttcct tcttcctcga tgccccggtg 120
gcgcgtctcc attgtctaag tggccacggt gctaagaaga aacgacttca ctttgacatt 180
caacagggaa gaaatgctat aagcaagact cacactccag acttatatgc caaacaggaa 240
tacagtgtgc cagagattgt gaagggtgac gacaaggaag aggaagtggt caagattaaa 300
gaacatgttg acatcattaa gtccatgttg agctcaatgg aggatggaga gattagcatt 360
tcagcttatg acactgcttg ggttgcactc atacaagaca ttcacaacaa tggtgctcct 420
caattcccat ctagcctgct ctggatagct gagaatcagc taccggacgg gtcatggggt 480
gacagtcgcg tatttttagc atttgatcga ataatcaata cgttagcttg tgttgttgct 540
ctcaagtcat ggaatgttca tcctgataaa tgcgaaagag gaatctcctt tttgaaagaa 600
aatataagca tgctagaaaa agacgattcc gagcacatgc ttgttggctt tgaatttggt 660
ttccctgtgt tgcttgacat ggctcgaaga ttaggcatcg atgttcctga tgattcgcca 720
ttcttgcaag agatctacgt ccagagagac ctgaaactca aaaggatacc aaaagacata 780
ttgcacaatg tgcccacgac actgctccat agcttggaag caataccaga cctggattgg 840
acaaagcttc taaaactaca atgtcaagat ggttcactct tgttctcccc atcctccact 900
gccatggcat tcatcaacac taaggacgaa aactgcttga gatatctaaa ttacgtggtc 960
cagagattca atggtggagc cccaacggtt tacccctacg acttgtttga gcacaattgg 1020
gcggttgatc gcctacaacg cttgggaatc tcaagatttt ttcagccaga gattagagaa 1080
tgtatgagct atgtttacag gtattggact aaagatggga tcttctgcac aagaaattca 1140
cgggttcatg atgttgacga tacagccatg ggattcaggt tgcttagatt gcacggctat 1200
gaagttcacc ctgatgcatt taggcagttt aagaagggat gcgagtttat atgctatgaa 1260
ggccaatcgc atccgacagt cactgtaatg tataacctgt acagggcttc tcagttgatg 1320
tttcctgaag aaaagattct tgacgaggcc aagcagttca cagaaaaatt tctgggagaa 1380
aaacgatcgg ctaataagct cctagataag tggatcataa ctaaggatct gccaggggag 1440
gtggggttcg cattggatgt tccgtggtat gcatccttgc ctagagtgga ggcaagattc 1500
ttcattcaac actacggcgg cgaagatgat gtatggcttg acaaggccct ttacaggatg 1560
ccctacgtta acaacaatgt gtatctagag ctagcaaagc ttgattacaa ctattgccaa 1620
gcattgcatc gaaccgaatg gggccgtata caaaagtggt atgaagaaag taaaccaaga 1680
gactttggga taagcagaga atgccttctc cgtgcatatt ttatggctgc tgccagtata 1740
tttgagccag aaaggtcgat ggagcgcctc gcctgggcta agactgcgat cttgctggaa 1800
attatagtat cttatttcaa tgaggttgga aattccacag agcagaggat agccttcaca 1860
actgaatttt cgatacgtgc aagtcctatg ggaggctaca taaatggaag aaagttagat 1920
aagattggta cgacccaaga acttattcag atgctactcg caaccatcga tcagttttcg 1980
caggatgcat tcgcggcata tggtcacgac attactcgcc atttacacaa cccttggaaa 2040
atgtggctgc taaagtggca agaagaagga gatagatggc tgggagaagc agagttactg 2100
atacaaacta taaatctcat ggcggatcat aagatcgctg agaagctctt tatgggacat 2160
actaactatg aacaactctt cagcctcacc aacaaagttt gctatagtct tggtcatcat 2220
gagctccaaa acaacagaga actggagcat gacatgcaaa gactagtgca gttggtgctg 2280
acgaattcct cagatggtat cgactctgat atcaagaaaa cgtttctcgc agtcgcaaag 2340
agattctact acactgcctt tgtcgaccct gagaccgtga atgtccacat tgccaaagta 2400
ctctttgaga gagtagatta a 2421
<210> 3
<211> 36
<212> DNA
<213> 人工序列
<400> 3
cgcggatcca tgcatagttt gttaatgaaa aaggtg 36
<210> 4
<211> 38
<212> DNA
<213> 人工序列
<400> 4
gcactgcagt taatctactc tctcaaagag tactttgg 38
雷公藤焦磷酸合酶TwCPS4及其制备松香烷型二萜化合物的应用专利购买费用说明
![]()
Q:办理专利转让的流程及所需资料
A:专利权人变更需要办理著录项目变更手续,有代理机构的,变更手续应当由代理机构办理。
1:专利变更应当使用专利局统一制作的“著录项目变更申报书”提出。
2:按规定缴纳著录项目变更手续费。
3:同时提交相关证明文件原件。
4:专利权转移的,变更后的专利权人委托新专利代理机构的,应当提交变更后的全体专利申请人签字或者盖章的委托书。
Q:专利著录项目变更费用如何缴交
A:(1)直接到国家知识产权局受理大厅收费窗口缴纳,(2)通过代办处缴纳,(3)通过邮局或者银行汇款,更多缴纳方式
Q:专利转让变更,多久能出结果
A:著录项目变更请求书递交后,一般1-2个月左右就会收到通知,国家知识产权局会下达《转让手续合格通知书》。
















动态评分
0.0