









专利摘要
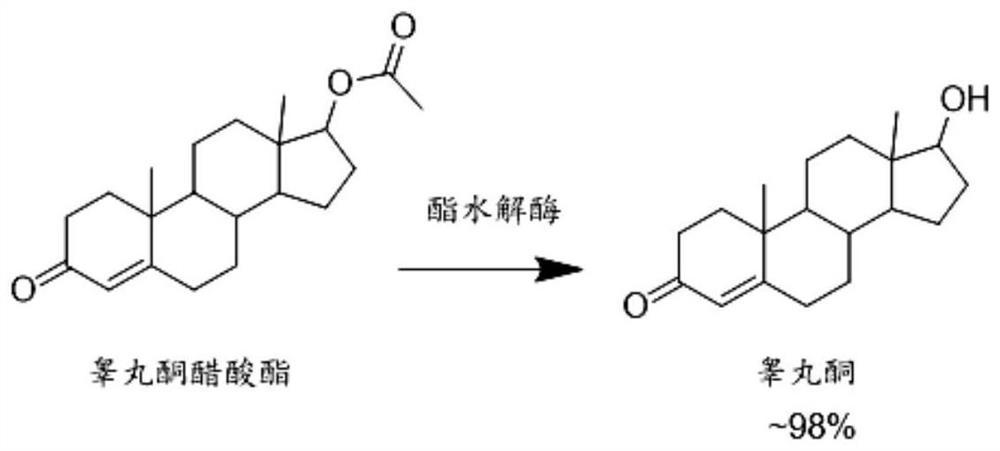
本发明公开了一种通过分子进化获得的活性和稳定性提高的7β‑羟基甾醇脱氢酶突变体、含有7β‑羟基甾醇脱氢酶突变基因的重组表达质粒及重组表达转化子、重组突变体酶制剂的制备方法,以及该重组突变体酶制剂在熊脱氧胆酸合成中的应用。本发明的7β‑羟基甾醇脱氢酶具有出色的活性和热稳定性,可以高效催化7‑羰基石胆酸不对称还原制备熊脱氧胆酸;将本发明所述的7β‑羟基甾醇脱氢酶固定化后与固定化7α‑羟基甾醇脱氢酶酶法耦联,可以直接催化廉价底物鹅脱氧胆酸的差向异构,连续转化制备熊脱氧胆酸,操作简便。与现有报道相比,本发明的羟基甾醇脱氢酶催化制备熊脱氧胆酸,具有底物浓度高、反应时间短、反应完全,产品纯度高的优势,有很强的工业应用前景。
权利要求
1.一种活性及稳定性提高的7β-羟基甾醇脱氢酶,其特征在于,其是将如序列表中SEQ ID No.2所示氨基酸序列的蛋白质的第57位甘氨酸、第128位甘氨酸、第163位异亮氨酸、第172位丙氨酸、第176位丝氨酸、第197位天冬酰胺、第200位甘氨酸、第243位天冬酰胺或第258位甘氨酸中的一个或多个氨基酸残基替换为其他氨基酸残基形成的新氨基酸序列的衍生蛋白质。
2.根据权利要求1所述的7β-羟基甾醇脱氢酶,其特征在于,所述7β-羟基甾醇脱氢酶具有如下序列:
(1)将如序列表中SEQ ID No.2所示氨基酸序列的第57位甘氨酸替换为苏氨酸,第128位甘氨酸替换为丙氨酸,同时第200位甘氨酸替换为天冬酰胺;
(2)将如序列表中SEQ ID No.2所示氨基酸序列的第57位甘氨酸替换为苏氨酸,第176位丝氨酸替换为丙氨酸,第258位甘氨酸替换为丙氨酸;
(3)将如序列表中SEQ ID No.2所示氨基酸序列的第57位甘氨酸替换为苏氨酸,第172位丙氨酸替换为亮氨酸;
(4)将如序列表中SEQ ID No.2所示氨基酸序列的第128位甘氨酸替换为甲硫氨酸,第163位异亮氨酸替换为缬氨酸;
(5)将如序列表中SEQ ID No.2所示氨基酸序列的第163位异亮氨酸替换为缬氨酸,第172位丙氨酸替换为异亮氨酸,第243位天冬酰胺替换为色氨酸;
(6)将如序列表中SEQ ID No.2所示氨基酸序列的第128位甘氨酸替换为甲硫氨酸,第197位天冬酰胺替换为苏氨酸,第258位甘氨酸替换为苏氨酸;
(7)将如序列表中SEQ ID No.2所示氨基酸序列的第172位丙氨酸替换为异亮氨酸,第197位天冬酰胺替换为酪氨酸;
(8)将如序列表中SEQ ID No.2所示氨基酸序列的第176位丝氨酸替换为亮氨酸,第200位甘氨酸替换为谷氨酸,第243位天冬酰胺替换为亮氨酸;
(9)将如序列表中SEQ ID No.2所示氨基酸序列的第197位天冬酰胺替换为酪氨酸,第258位甘氨酸替换为丙氨酸;
(10)将如序列表中SEQ ID No.2所示氨基酸序列的第57位甘氨酸替换为苏氨酸,第176位丝氨酸替换为丙氨酸,第128位甘氨酸替换为甲硫氨酸,第197位天冬酰胺替换为苏氨酸,同时第258位甘氨酸替换为苏氨酸;
(11)将如序列表中SEQ ID No.2所示氨基酸序列的第57位甘氨酸替换为苏氨酸,第128位甘氨酸替换为丙氨酸,第172位丙氨酸替换为异亮氨酸,第197位天冬酰胺替换为酪氨酸,同时第200位甘氨酸替换为天冬酰胺;
(12)将如序列表中SEQ ID No.2所示氨基酸序列的第163位异亮氨酸替换为缬氨酸,第172位丙氨酸替换为异亮氨酸,第197位天冬酰胺替换为酪氨酸,第243位天冬酰胺替换为色氨酸。
3.一种分离的核酸,其特征在于,所述的核酸编码如权利要求1或2中任一项所述的7β-羟基甾醇脱氢酶。
4.一种重组表达载体,其特征在于,所述重组表达载体包含如权利要求3所述的核酸。
5.一种重组表达转化体,其特征在于,所述重组表达转化体包含如权利要求4所述的重组表达载体。
6.如权利要求5所述的重组表达转化体,其特征在于,所述重组表达转化体的宿主选择大肠杆菌或毕赤酵母。
7.一种重组7β-羟基甾醇脱氢酶催化剂,其特征在于,所述的重组7β-羟基甾醇脱氢酶催化剂是以下形式中的任意一种:
(1)培养如权利要求6所述的重组表达转化体,分离含有所述7β-羟基甾醇脱氢酶的转化体细胞;
(2)培养如权利要求6所述的重组表达转化体,分离含有所述7β-羟基甾醇脱氢酶的粗酶液;
(3)对所述7β-羟基甾醇脱氢酶进行固定化,制备的固定化7β-羟基甾醇脱氢酶。
8.一种酶法转化制备熊脱氧胆酸的方法,其特征在于,使用如权利要求7所述的重组7β-羟基甾醇脱氢酶催化剂催化7-羰基石胆酸的不对称还原反应,制备熊脱氧胆酸。
9.如权利要求8所述酶法转化制备熊脱氧胆酸的方法,其特征在于,酶法还原反应使用NADPH作为辅酶,反应液中加入葡萄糖脱氢酶催化葡萄糖氧化,实现辅酶NADPH的再生。
10.如权利要求8所述酶法转化制备熊脱氧胆酸的方法,其特征在于,所述的7-羰基石胆酸由7α-羟基甾醇脱氢酶催化鹅脱氧胆酸立体选择性氧化制备。
11.如权利要求8或10所述酶法转化制备熊脱氧胆酸的方法,其特征在于,将固定化7α-羟基甾醇脱氢酶催化鹅脱氧胆酸立体选择性氧化制备7-羰基石胆酸的反应与如权利要求7所述的重组7β-羟基甾醇脱氢酶催化剂催化7-羰基石胆酸的不对称还原反应耦联,以鹅脱氧胆酸为底物,连续转化制备熊脱氧胆酸。
12.如权利要求8所述酶法转化制备熊脱氧胆酸的方法,其特征在于,所述熊脱氧胆酸的分离、精制步骤为:将分离除去催化剂的反应液酸化,使用有机溶剂萃取,萃取液洗涤、干燥后浓缩结晶,获得精制的纯品。
说明书
技术领域
本发明属于生物工程技术领域,具体涉及活性和稳定性提高的扭链瘤胃球菌7β-羟基甾醇脱氢酶(7β-hydroxysteroid dehydrogenase,7β-HSDHRt)突变体、其重组表达载体以及重组表达转化子,重组7β-羟基甾醇脱氢酶催化剂的制备方法,以及重组7β-羟基甾醇脱氢酶催化剂在熊脱氧胆酸合成中的应用。
背景技术
熊胆汁制品具有很高的药用价值,在现代医学中通常用于治疗反流性胃炎、胆结石、胆源性胰腺炎、脂肪性肝病、药物性肝炎以及病毒性肝炎等胆汁淤积性肝病。熊脱氧胆酸(简写为UDCA)是熊胆汁制品中的重要有效成分,具有很高的药用价值,可改善慢性肝病的损害,在急性和慢性肝炎治疗中的应用正在扩大。UDCA在熊的胆汁中含量最高,而在其他动物的胆汁中含量很少。目前UDCA大部分是从人工养殖的熊的胆汁中提取,产量低,来源有限,并且直接从活熊身上抽取胆汁有悖于动物保护。如果以其他动物胆汁来源的,可大量获得的胆酸(CA)或者UDCA的差向异构体鹅脱氧胆酸(CDCA)作为底物,采用酶法或者化学-酶结合的方法,人工合成具有较高价值的UDCA,减少对天然熊胆汁的需求,将具有重要的经济、社会价值和生态意义。
早期研究中,以牛、羊胆汁来源的CA为原料,经甲酯化、二乙酰化、12位羟基氧化、黄鸣龙还原合成CDCA,进而氧化生成7-羰基石胆酸(7-KLCA),最后在正丙醇中用金属钠还原得到UDCA。这种方法步骤繁琐,操作安全性差,产品得率低,并且需要使用重金属氧化剂和催化剂,目前已被淘汰。
日本专利(JP 02282393)报道,在丁醇溶液中,氢氧化钠和钯碳存在的碱性条件下,于100℃,80kg/cm2的压力环境中,化学法催化7-KLCA的氢化反应,反应5h,UDCA的得率为88.2%。由于此方法需要在高压下反应,操作不便,没有投入实际应用。
2009年,Riva等报道了生物催化CA转化生成12-酮基-UDCA,进而利用化学法催化还原制备UDCA的方法(Adv Synth Catal,2009,351:1303-1311)。该生物转化反应中使用了三种酶,首先应用7α-羟基甾醇脱氢酶(7α-HSDH)和12α-羟基甾醇脱氢酶(12α-HSDH)催化CA氧化生成7,12-二酮基石胆酸,再利用7β-羟基甾醇脱氢酶(7β-HSDH)催化7,12-二酮基石胆酸还原生成12-酮基熊脱氧胆酸。随后通过wolff-kishner化学还原反应生成UDCA。由于该酶法氧化还原过程中催化辅酶循环再生的酶专一性差,导致转化不完全,因此终产物UDCA的化学纯度不高。
2011年,Schmid等公布了来自Collinsella aerofaciens的7β-HSDH基因序列,将其克隆表达于大肠杆菌,并利用该酶催化7-KLCA转化为UDCA,底物浓度40g/L,转化24h,转化率90%(Appl Microbiol Biotechnol,2011,90:127–135)。Juste等通过筛选,分离得到一些可以催化CDCA的7-位异构化,产生UDCA的野生型菌株(FEMS Microbiol Lett,2004,235:65–72),在此基础上,Monti等从Clostridium absonum中克隆获得另一个7β-HSDH,同时克隆获得一个7α-HSDH,这两个酶可联合作用,催化CDCA的差向异构制备UDCA(Appl Microbiol Biotechnol,2012,95:1221–1233)。2013年Yokota等从自然界筛选得到的野生菌Ruminococcus gnavus N53中克隆获得7β-HSDH,使用生长细胞催化7-KLCA转化制备UDCA,底物浓度1mM,转化率为90~100%(J Lipid Res,2013,54:3062-3069)。南京普瑞特生物科技有限公司对来源于Ruminococcus gnavus的7β-HSDH进行了活性改造,并与醇脱氢酶偶联构建辅酶再生循环,催化7KLCA转化制备UDCA,底物浓度达100g/L,转化率>99%。苏州汉酶生物技术有限公司使用来源于Ruminococcaceae bacterium的7-KLCA还原酶及其突变体作为催化剂,催化7-KLCA还原制备UDCA,底物浓度100g/L,4h反应完毕。华东理工大学对Ruminococcus torques中克隆得到的7β-HSDHRt进行了活性改造,并与来源于Clostridium absonum的7α-HSDHCa以及相应的辅酶再生酶相耦联,一锅催化CDCA差向异构制备UDCA,底物浓度为100mM时,最终转化率高于99%(Process Biochem,2015,50:598-604,J Agric Food Chem,2017,65:1178-1185),但该报道中,由于辅酶再生用酶葡萄糖脱氢酶底物专一性差,因此一锅反应分两步进行,待第一步7α-HSDHCa催化的CDCA氧化反应结束后,需要对酶进行灭活处理,再加入7β-HSDHRt以及葡萄糖脱氢酶,催化中间产物7-KLCA的不对称还原,制备获得UDCA。
综上所述,现有的酶法制备UDCA的报道中,酶法催化7-KLCA不对称还原制备UDCA的报道较多,但所使用的7β-HSDH活性偏低,并且都使用大肠杆菌作为重组表达的宿主,表达的酶是胞内酶,需要对重组菌进行细胞破碎,才能分离获得酶液,应用过程繁琐,并且7-KLCA并非自然界可大量提取的动物胆酸类原料,价格较高;而CDCA虽然在家禽胆汁中大量存在,价格相对低廉,但酶法耦联催化CDCA差向异构的报道较少,底物浓度不高,并且已报道的两步酶法反应中,需要对第一步反应的酶进行灭活处理,才能加入第二步反应的酶,步骤繁琐,效率低下,酶不能重复应用,生物催化剂应用成本较高。
发明内容
本发明的目的在于,在已报道的7β-HSDHRt突变体的基础上,通过蛋白质工程和定向进化的手段对其进行进一步改造,进一步提高该酶的活性和热稳定性;选择毕赤酵母作为表达宿主,对高活性和稳定性的重组7β-HSDHRt突变体进行胞外表达,简化酶的分离工艺;对重组表达的7β-HSDHRt突变体进行固定化,与同样经过固定化的7α-HSDHCa进行高效酶促反应耦联,实现双酶反应的空间分离,从而连续高效催化CDCA差向异构化合成UDCA,有效简化UDCA的合成工艺,降低酶催化剂的应用成本。
本发明的目的可以通过以下技术方案来实现:
本发明的技术方案之一:获得活性和热稳定性提高的7β-HSDHRt突变体。在先前报道中,通过对7β-HSDHRt的蛋白质工程改造,获得了如序列表SEQ ID No.2所示氨基酸序列的羟基甾醇脱氢酶7β-HSDHRt-T89V/V207M。本发明中,以该酶基因作为母本,采用易错PCR、迭代饱和突变、DNA shuffling等组合突变策略对其进行进一步定向进化,结合酶标仪高通量初筛和自动进样HPLC复筛,鉴别获得活性和热稳定性显著改善的7β-羟基甾醇脱氢酶。
所述活性及稳定性提高的7β-羟基甾醇脱氢酶(7β-HSDH),是将如序列表中SEQ ID No.2所示氨基酸序列的蛋白质的第57位甘氨酸、第128位甘氨酸、第163位异亮氨酸、第172位丙氨酸、第176位丝氨酸、第197位天冬酰胺、第200位甘氨酸、第243位天冬酰胺或第258位甘氨酸中的一个或多个氨基酸残基替换为其他氨基酸残基形成的新氨基酸序列的衍生蛋白质。
具体的,所述7β-羟基甾醇脱氢酶具有如下序列:
(1)将如序列表中SEQ ID No.2所示氨基酸序列的第57位甘氨酸替换为苏氨酸,第128位甘氨酸替换为丙氨酸,同时第200位甘氨酸替换为天冬酰胺,将该酶命名为7β-M1;
(2)将如序列表中SEQ ID No.2所示氨基酸序列的第57位甘氨酸替换为苏氨酸,第176位丝氨酸替换为丙氨酸,第258位甘氨酸替换为丙氨酸,将该酶命名为7β-M2;
(3)将如序列表中SEQ ID No.2所示氨基酸序列的第128位甘氨酸替换为甲硫氨酸,第163位异亮氨酸替换为缬氨酸,将该酶命名为7β-M3;
(4)将如序列表中SEQ ID No.2所示氨基酸序列的第163位异亮氨酸替换为缬氨酸,第172位丙氨酸替换为异亮氨酸,第243位天冬酰胺替换为色氨酸,将该酶命名为7β-M4;
(5)将如序列表中SEQ ID No.2所示氨基酸序列的第128位甘氨酸替换为甲硫氨酸,第197位天冬酰胺替换为苏氨酸,第258位甘氨酸替换为苏氨酸,将该酶命名为7β-M5;
(6)将如序列表中SEQ ID No.2所示氨基酸序列的第57位甘氨酸替换为苏氨酸,第172位丙氨酸替换为亮氨酸,将该酶命名为7β-M6;
(7)将如序列表中SEQ ID No.2所示氨基酸序列的第172位丙氨酸替换为异亮氨酸,第197位天冬酰胺替换为酪氨酸,将该酶命名为7β-M7;
(8)将如序列表中SEQ ID No.2所示氨基酸序列的第176位丝氨酸替换为亮氨酸,第200位甘氨酸替换为谷氨酸,第243位天冬酰胺替换为亮氨酸,将该酶命名为7β-M8;
(9)将如序列表中SEQ ID No.2所示氨基酸序列的第197位天冬酰胺替换为酪氨酸,第258位甘氨酸替换为丙氨酸,将该酶命名为7β-M9;
(10)将如序列表中SEQ ID No.2所示氨基酸序列的第57位甘氨酸替换为苏氨酸,第176位丝氨酸替换为丙氨酸,第128位甘氨酸替换为甲硫氨酸,第197位天冬酰胺替换为苏氨酸,同时第258位甘氨酸替换为苏氨酸,将该酶命名为7β-M10;
(11)将如序列表中SEQ ID No.2所示氨基酸序列的第57位甘氨酸替换为苏氨酸,第128位甘氨酸替换为丙氨酸,第172位丙氨酸替换为异亮氨酸,第197位天冬酰胺替换为酪氨酸,同时第200位甘氨酸替换为天冬酰胺,将该酶命名为7β-M11;
(12)将如序列表中SEQ ID No.2所示氨基酸序列的第163位异亮氨酸替换为缬氨酸,第172位丙氨酸替换为异亮氨酸,第197位天冬酰胺替换为酪氨酸,第243位天冬酰胺替换为色氨酸,将该酶命名为7β-M12。
所述7β-羟基甾醇脱氢酶的活力测定方法:将含2mmol/L 7-KLCA和0.1mmol/L NADPH的1ml反应体系(100mmol/L磷酸钾缓冲液,pH 8.0)预热至30℃,然后加入适量的羟基甾醇脱氢酶,30℃保温反应,在分光光度计上检测340nm处NADPH的吸光度变化,记录1分钟内吸光度的变化值。
用下式计算得到酶活力:
酶活力(U)=EW×V×103/(6220×l)
式中,EW为1分钟内340nm处吸光度的变化;V为反应液的体积,单位为ml;6220为NADPH的摩尔消光系数,单位为L/(mol·cm);l为光程距离,单位为cm。1个活力单位(U)对应于上述条件下每分钟催化氧化1μmol NADPH所需的酶量。
本发明的技术方案之二:一种分离的核酸,以及含有所述核酸分子的重组表达载体。所述核酸编码如技术方案一所述的7β-羟基甾醇脱氢酶,其来源包括:通过基因工程技术对技术方案一所述的进化改造获得的系列7β-羟基甾醇脱氢酶的基因序列进行克隆;或者通过人工全序列合成的方法得到编码如技术方案一所述7β-羟基甾醇脱氢酶的核酸分子。
所述的重组表达载体可通过本领域常规方法,将本发明所述的7β-羟基甾醇脱氢酶的编码核酸分子连接于各种载体上构建而成。所述的载体可以是本领域常规的各种质粒载体,只要所述重组表达载体可以在相应的表达宿主中正常复制,并表达相应的7β-羟基甾醇脱氢酶即可。针对不同的表达宿主,优选的质粒载体是不同的。对于大肠杆菌宿主,所述质粒载体优选的为pET28a质粒;对于毕赤酵母宿主,所述质粒载体优选的为pPICZαA。
以含有所述羟基甾醇脱氢酶7β-M6编码核酸分子的重组表达载体的制备为例,较佳的,可通过下述方法制得本发明所述的大肠杆菌重组表达载体pET28a-7β-M6:
设计如下引物对所述7β-羟基甾醇脱氢酶突变体基因的DNA序列进行扩增:
上游引物,如序列表SEQ ID No.3所示:
5’–CCGGAATTCATGAATCTGCGTGAAAAATAC-3’,下划线所示序列为限制性内切酶EcoR I的酶切位点;
下游引物,如序列表SEQ ID No.4所示:
5’–CCGCTCGAGTTAATTGTTGCTATAGAAGC-3’,下划线所示序列为限制性内切酶Xho I的酶切位点。
将通过PCR扩增所得的7β-羟基甾醇脱氢酶基因序列DNA片段用限制性内切酶EcoR I和Xho I双酶切,同时将空载质粒pET28a同样用限制性内切酶EcoR I和Xho I双酶切,回收上述酶切后的7β-羟基甾醇脱氢酶基因DNA片段以及空载质粒,利用T4 DNA连接酶连接,构建获得用于大肠杆菌表达的含有所述7β-羟基甾醇脱氢酶编码核酸分子的重组表达载体pET28a-7β-M6。
采用类似的方法和技术,可以方便地构建获得用于毕赤酵母表达的含有所述7β-羟基甾醇脱氢酶编码核酸分子的重组表达载体pPICZαA-7β-M6。
本发明的技术方案之三:包含所述7β-羟基甾醇脱氢酶重组表达载体的重组表达转化体。所述重组表达转化体可通过本领域常规技术,将上述重组表达载体转化至相应宿主细胞中制得。所述宿主细胞为本领域常规的宿主细胞,只要能满足重组表达载体可稳定地自行复制,并且其所编码的7β-羟基甾醇脱氢酶基因可被有效表达即可。所述宿主细胞优选大肠杆菌和毕赤酵母,更优选地为:大肠杆菌E.coli BL21(DE3)和毕赤酵母P.pastoris X33。
采用本领域常规技术,将所述重组表达载体分别转化至E.coli BL21(DE3)或毕赤酵母P.pastoris X33中,即可获得本发明优选的重组表达转化体基因工程菌株。例如,将重组表达载体pET28a-7β-M6转化至大肠杆菌E.coli BL21(DE3)中,即可获得重组大肠杆菌E.coli BL21(DE3)/pET28a-7β-M6;将重组表达载体pPICZαA-7β-M6转化至毕赤酵母P.pastoris X33中,即可获得重组毕赤酵母P.pastoris X33/pPICZαA-7β-M6。
本发明的技术方案之四:7β-羟基甾醇脱氢酶的表达以及酶催化剂的制备方法。
所述重组表达转化体的培养条件为本领域常规的方法和条件,针对使用不同宿主构建的重组表达转化体,采用不同的优选培养条件。
对于重组大肠杆菌,摇瓶培养使用的优选培养基为LB培养基:蛋白胨10g/L,酵母膏5g/L,NaCl 10g/L,pH 7.0。培养方法和培养条件没有特殊的限制,只要所述重组大肠杆菌能够生长并高效表达所述的羟基甾醇脱氢酶即可。对于重组E.coli BL21(DE3),优选的培养方法为:将如上所述构建的重组大肠杆菌,接种至含有硫酸卡那霉素的LB培养基中,37℃、180rpm振荡培养过夜。按1~2%(v/v)的接种量接入装有100ml的LB培养基(含有硫酸卡那霉素)的500ml三角烧瓶中,置于37℃、180rpm摇床振荡培养,当培养液的OD600达到0.6~0.8时,加入终浓度为0.1~0.5mmol/L的异丙基-β-D-硫代半乳糖苷(IPTG)作为诱导剂,16~25℃诱导16~24h后,将培养液离心,收集沉淀,然后用生理盐水洗涤两次,获得重组表达转化体细胞。将收获的重组表达转化体进行冷冻干燥,即可获得含有所述7β-羟基甾醇脱氢酶的冻干细胞。将离心收获的重组表达转化体细胞悬浮于5~10倍体积(v/w)的缓冲液中,超声破碎,离心收集上清液,即可获得所述重组7β-羟基甾醇脱氢酶的粗酶液。
对于重组毕赤酵母菌,摇瓶培养使用的培养基优选BMGY培养基:甘油10g/L,蛋白胨20g/L,酵母提取物10g/L,生物素40mg/L,无氨基酸酵母氮源13.4g/L以及终浓度为100mM的磷酸钾缓冲盐,pH 6.0。培养方法和培养条件没有特殊的限制,只要所述重组毕赤酵母能够生长并高效表达所述的7β-羟基甾醇脱氢酶即可。对于重组毕赤酵母X33宿主构建的重组表达转化体,优选下述方法:将表达本发明所述重组7β-羟基甾醇脱氢酶的重组酵母菌接种至含氨苄青霉素的BMGY培养基中培养,培养温度为20~30℃。当培养液的光密度OD600达到1.3~2(优选1.5)时,将培养基替换为BMMY(甲醇10ml/L,蛋白胨20g/L,酵母提取物10g/L,生物素40mg/L,无氨基酸酵母氮源13.4g/L以及终浓度为100mM的磷酸钾缓冲盐,pH 6.0),每隔24h添加相当于培养液体积1%的纯甲醇进行诱导,持续诱导96h,可高效诱导重组毕赤酵母菌分泌表达本发明的重组7β-羟基甾醇脱氢酶。培养结束后,将培养液高速离心,收集离心上清液,同样可获得所述7β-羟基甾醇脱氢酶的粗酶液。
将含有所述7β-羟基甾醇脱氢酶的粗酶液用缓冲液适当稀释,加入预处理的环氧树脂或氨基化树脂,酶蛋白与树脂的比例为1~50mg酶蛋白/g树脂,4~30℃振摇,固定化1~24h,然后用缓冲液洗涤,获得固定化7β-羟基甾醇脱氢酶。所述缓冲液为本领域常规的缓冲溶液,优选磷酸盐缓冲液,pH 5.5~8.5。按照类似的方法,可以将7β-羟基甾醇脱氢酶与葡萄糖脱氢酶的共同固定化于树脂载体上,获得7β-羟基甾醇脱氢酶-葡萄糖脱氢酶的共固定化催化剂;类似的,可以将羟基甾醇脱氢酶7α-HSDHCa与乳酸脱氢酶共同固定化于树脂载体上,获得7α-HSDHCa-乳酸脱氢酶的共固定化催化剂。
本发明的技术方案之五:所述7β-羟基甾醇脱氢酶催化剂在熊脱氧胆酸合成中的应用。
所述应用方案包括:(1)以7-KLCA作为底物,使用上述制备的7β-羟基甾醇脱氢酶催化剂催化7-KLCA不对称还原,直接制备获得UDCA;
或者(2)以CDCA作为底物,将上述制备的7β-羟基甾醇脱氢酶-葡萄糖脱氢酶共固定化催化剂与7α-HSDHCa-乳酸脱氢酶共固定化催化剂进行酶法耦联催化CDCA的差向异构,制备获得UDCA。
所述7β-羟基甾醇脱氢酶突变体催化剂催化7-KLCA不对称还原,制备UDCA的方法,可按下述方法进行:在pH 6.0~9.0的缓冲盐溶液中,在葡萄糖脱氢酶、葡萄糖和额外添加的NADP+的存在下,加入7-KLCA和如上所述的7β-羟基甾醇脱氢酶催化剂,恒温、充分混合的条件下,酶法催化7-KLCA的不对称还原反应。所述应用中,底物7-KLCA的浓度为10~120g/L。7β-羟基甾醇脱氢酶催化7-KLCA还原时,辅酶NADPH氧化生成NADP+,为了进行辅酶NADPH的循环再生,向反应体系中额外添加葡萄糖和来自巨大芽孢杆菌的葡萄糖脱氢酶(中国现代医学杂志,2007,17(2):172-174)。葡萄糖脱氢酶的活力单位上载可以与所述7β-羟基甾醇脱氢酶相等。葡萄糖与底物的摩尔比可以为1.0~1.5,额外添加的辅酶NADP+的用量可以为0~1.0mmol/L。所述缓冲盐溶液可以是本领域常规的任何缓冲液,只要其pH范围在6.0~9.0即可,比如磷酸钠、磷酸钾、Tris-HCl或者甘氨酸-NaOH缓冲液,优选pH范围为7.0~8.0,更优选pH 8.0的磷酸钾缓冲液。缓冲液的浓度可以为0.05~0.2mol/L。所述的酶促不对称还原反应的温度可以是25~40℃,优选35℃。反应过程中,间歇取样测定反应转化率,反应时间以底物完全转化或反应转化率停止增长的时间为准,一般为0.5~24小时。反应转化率采用液相色谱法进行分析,使用C-18柱(250mm×4.6mm),流动相为甲醇/水=75:25(磷酸调节pH为3.0),柱温30℃,流速0.8ml/min,检测波长210nm。
所述7β-羟基甾醇脱氢酶-葡萄糖脱氢酶共固定化催化剂与7α-HSDHCa-乳酸脱氢酶进行酶法耦联催化CDCA的差向异构,制备获得UDCA的方法,优选的,可按下述方法进行:按技术方案之四所述的方法,将所述的7β-羟基甾醇脱氢酶与葡萄糖脱氢酶共固定化于环氧树脂ES-103上,同时将7α-HSDHCa与乳酸脱氢酶共固定化于环氧树脂ES-103上,将所述的7β-羟基甾醇脱氢酶-葡萄糖脱氢酶的共固定化催化剂7β-M6/GDH@ES-103和7α-HSDHCa-乳酸脱氢酶共固定化催化剂7α-HSDH/LDH@ES-103分别装柱,将两根柱串联,使含有底物CDCA、辅底物(葡萄糖和丙酮酸)以及辅酶(NAD+和NADP+)的缓冲溶液依次通过7α-HSDH/LDH@ES-103和7β-HSDH-M6/GDH@ES-103的酶柱,即可实现连续反应,一步法直接获得UDCA。所述缓冲盐溶液可以是本领域常规的任何缓冲液,只要其pH范围在6.0~9.0即可,比如磷酸钠、磷酸钾、Tris-HCl或者甘氨酸-NaOH缓冲液,优选pH范围为7.0~8.0,更优选pH 8.0的磷酸钾缓冲液。磷酸盐缓冲液的浓度可以为0.05~0.2mol/L。所述底物CDCA的浓度为10~120g/L,额外添加辅酶NAD+和NADP+的浓度为0.05~0.5mmol/L,所述的酶促不对称还原反应的温度可以是25~40℃,优选35℃。在优化条件下,平衡转化率高于99%。
本发明的技术方案之六:一种产物UDCA的提取方法。由于本发明的7-KLCA不对称还原反应以及CDCA的差向异构反应具有很高的转化率,反应液中只有极低浓度的7-KLCA残余。反应结束后,将催化剂分离除去,然后将反应液酸化,用常规溶剂萃取,萃取液洗涤、干燥后浓缩,即可结晶获得高纯度的UDCA。
与现有技术相比,本发明的7β-羟基甾醇脱氢酶具有更高的催化活性和热稳定性,催化反应底物浓度更高,转化完全,反应的时空产率高;使用毕赤酵母宿主体系分泌表达本发明所述的7β-羟基甾醇脱氢酶,与文献报道的大肠杆菌宿主体系相比,酶的分离提取更简便;使用固定化酶耦联催化CDCA差向异构制备UDCA,克服了文献中需对第一步反应的酶进行灭活处理的问题,可以连续转化,操作简便,酶催化剂可长期重复使用,具有很强的工业应用潜力。此外,本发明的所有试剂和原料均市售可得,实用性强。
具体实施方式
下面结合具体实施例对本发明进行详细说明。
发明内容中所述的各反应或检测条件,可根据本领域常识进行组合或更改,并可通过实验得到验证。下面通过实施例的方式进一步说明本发明。
下列实施例中的材料来源为:
质粒pET28a-7β-M0,含有如序列表SEQ ID No.1所示的核酸序列,为申请人自行构建(参见:J Agric Food Chem,2017,65:1178-1185)。
空载质粒pET28a和pPICZαA购自Novagen公司。
E.coli DH5α、E.coli BL21(DE3)和毕赤酵母X33感受态细胞、2×Taq PCR MasterMix、琼脂糖凝胶DNA回收试剂盒均购自北京天根生化科技有限公司。
限制性内切酶EcoR I和Xho I均为New England Biolabs(NEB)公司的市售产品。
环氧树脂ES-103购自天津南开和成科技有限公司。
除非另有说明,下列实施例中的具体实验按照本领域常规方法和条件进行,或遵照试剂盒的商品说明书。
实施例1 随机突变筛选活性与稳定性提高的7β-羟基甾醇脱氢酶
利用易错PCR的方法对如序列表中SEQ ID No.2所示氨基酸序列的7β-羟基甾醇脱氢酶进行随机突变。
使用的引物为:
上游引物,如序列表SEQ ID No.3所示:
5’–CCGGAATTCATGAATCTGCGTGAAAAATAC-3’,其中下划线所示序列为限制性内切酶EcoR I的酶切位点;
下游引物,如序列表SEQ ID No.4所示:
5’–CCGCTCGAGTTAATTGTTGCTATAGAAGC-3’,其中下划线所示序列为限制性内切酶Xho I的酶切位点。
PCR体系(50μL):Taq mix 25μl,终浓度为100μmol/L的MnCl2,pET28a-7β-M0质粒约1ng,上下游引物各2μl,diH2O补足至50μl。
PCR反应程序:(1)95℃预变性5min;(2)94℃变性30s;(3)58℃退火30s;(4)72℃延伸1min;(5)步骤(2)~(4)共进行30个循环;(6)72℃延伸10min,最后4℃保存产物。
将得到的PCR产物DNA片段与空载质粒pET28a分别用限制性内切酶EcoR I和Xho I进行双酶切。体系如下:
37℃酶切3h,切胶回收酶切后的目的片段和质粒。将回收纯化后的PCR片段与pET28a在16℃连接过夜。体系如下:
取10μL连接产物用CaCl2法转化50μL大肠杆菌BL21(DE3)感受态细胞,并均匀涂布含有50μg/ml硫酸卡那霉素的LB琼脂平板,37℃培养12h。
在96孔深孔板的每个孔洞中分别加入300μL无菌的LB培养基(含有50μg/ml硫酸卡那霉素)。将转化平板上的转化体用牙签挑入96孔深孔板中,于37℃,220rpm摇床中培养过夜。从一级板的孔洞中吸取50μL菌液接入二级板的相应孔洞中,于37℃,220rpm摇床中培养2~3h后,加入终浓度为0.2mM的IPTG,16℃培养20h。然后于4℃、3500×g离心10min,弃去培养基,放入-80℃冰箱冷冻2h。从冰箱中拿出深孔板,待菌液融化后,每个孔洞中加入200μL溶菌酶液(750mg溶菌酶和10mg DNA酶溶解于1L去离子水中),振荡混匀,37℃静置1h。4℃、3500×g离心10min,将细胞破碎离心上清液转移到新的96孔板中,用于后续活性和稳定性筛选。
突变体的活性筛选:在96孔板中进行,每个样孔加150μL反应液(100mM KPB,pH 8.0,含有1mM 7-KLCA以及0.2mM NADPH),随后加入50μL酶液,30℃、震荡混匀,在酶标仪上读取340nm处吸光度值的减少。
突变体的热稳定性筛选:在96孔板中进行,每个样孔中加入50μL活性筛选相对应的酶液,在45℃保温15min后,加入150μL反应液(100mM KPB,pH 8.0,含有1mM 7-KLCA以及0.2mM NADPH),震荡混匀,在酶标仪上读取340nm处吸光度值的减少,计算酶的残余活力。
通过筛选,得到活性和热稳定性均有明显提高的克隆,对相应的酶进行序列测定。表1提供了易错PCR突变库中筛选获得的突变体的序列以及相应的活性和稳定性列表。其中,突变体编号分别对应于表1后面列的一些序列。在活性列中,一个加号“+”表示突变体蛋白的比活力比由序列表中SEQ ID No.2所示氨基酸序列组成的蛋白质提高了0.1~1.0倍;两个加号“++”表示突变体蛋白的比活力比SEQ ID No.2所示氨基酸序列组成的蛋白质提高了1.1~2.0倍;三个加号“+++”表示突变体蛋白的比活力比SEQ ID No.2所示氨基酸序列组成的蛋白质提高了2.1~3.0倍。在热稳定性列中,一个加号“+”对应于45℃保温15min后,突变体蛋白的残余活性保留30.0~45.0%;两个加号“++”对应于45℃保温15min后,突变体蛋白的残余活性保留45.1~60.0%;三个加号“+++”对应于45℃保温15min后,突变体蛋白的残余活性保留60.1~80.0%。
如序列表中SEQ ID No.2所示氨基酸序列的7β-羟基甾醇脱氢酶的比活为17.6U/mg蛋白,45℃保温15min后,酶的残余活性为32.1%。
表1:7β-羟基甾醇脱氢酶序列和相应的活性改进列表
对应突变体编号的7β-羟基甾醇脱氢酶的氨基酸序列分别如下:
M1:将如序列表中SEQ ID No.2所示氨基酸序列的第57位甘氨酸替换为苏氨酸,第128位甘氨酸替换为丙氨酸,第200位甘氨酸替换为天冬酰胺;
M2:将如序列表中SEQ ID No.2所示氨基酸序列的第57位甘氨酸替换为苏氨酸,第176位丝氨酸替换为丙氨酸,第258位甘氨酸替换为丙氨酸;
M3:将如序列表中SEQ ID No.2所示氨基酸序列的第128位甘氨酸替换为甲硫氨酸,第163位异亮氨酸替换为缬氨酸;
M4:将如序列表中SEQ ID No.2所示氨基酸序列的第163位异亮氨酸替换为缬氨酸,第172位丙氨酸替换为异亮氨酸,第243位天冬酰胺替换为色氨酸;
M5:将如序列表中SEQ ID No.2所示氨基酸序列的第128位甘氨酸替换为甲硫氨酸,第197位天冬酰胺替换为苏氨酸,第258位甘氨酸替换为苏氨酸;
M6:将如序列表中SEQ ID No.2所示氨基酸序列的第57位甘氨酸替换为苏氨酸,第172位丙氨酸替换为亮氨酸;
M7:将如序列表中SEQ ID No.2所示氨基酸序列的第172位丙氨酸替换为异亮氨酸,第197位天冬酰胺替换为酪氨酸;
M8:将如序列表中SEQ ID No.2所示氨基酸序列的第176位丝氨酸替换为亮氨酸,第200位甘氨酸替换为谷氨酸,第243位天冬酰胺替换为亮氨酸;
M9:将如序列表中SEQ ID No.2所示氨基酸序列的第197位天冬酰胺替换为酪氨酸,第258位甘氨酸替换为丙氨酸;
实施例2 7β-羟基甾醇脱氢酶的组合突变
按实施例1所述的方法以及引物,对实施例1获得的7β-羟基甾醇脱氢酶M1-M9的基因DNA序列进行PCR扩增,将获得的DNA片段等比例混合,加入DNase I进行消化,反应体系200μl,含有100mM Tris-HCl(pH 7.5)、0.1U的DNase I、20μg的混合DNA片段和10mM MnCl2,37℃酶切3分钟,电泳分离,收集50~200bp的酶切片段,进行PCR组装。
PCR组装反应体系50μl,含有反应缓冲液,100ng的酶切片段、0.2mM dNTP mix和1μl的KOD高保真聚合酶。
PCR反应程序:(1)95℃预变性2min;(2)94℃变性30s;(3)65℃退火30s;(4)60℃退火30s;(5)55℃退火30s;(6)50℃退火30s;(7)45℃退火30s;(8)68℃延伸1min;(9)步骤(2)~(8)共进行45个循环;(10)68℃延伸5min。PCR产物稀释200倍,按实施例1所述方法,使用如序列表SEQ ID No.3和4所示序列的上下游引物,对PCR组装产物进行扩增,并连接到空载质粒pET28a上,然后转化大肠杆菌BL21(DE3)感受态细胞,并进行活性和稳定性筛选。
表2提供了组合突变库中筛选获得的突变体的序列以及相应的活性和稳定性列表。其中,突变体编号分别对应于表2后面列的一些序列,在活性列中,一个加号“+”表示突变体蛋白的比活力比由序列表中SEQ ID No.2所示氨基酸序列组成的蛋白质提高了0.1~1.0倍;两个加号“++”表示突变体蛋白的比活力比SEQ ID No.2所示氨基酸序列组成的蛋白质提高了1.1~2.0倍;三个加号“+++”表示突变体蛋白的比活力比SEQ ID No.2所示氨基酸序列组成的蛋白质提高了2.1~3.0倍。在热稳定性列中,一个加号“+”对应于45℃保温15min后,突变体蛋白的残余活性保留30.0~45.0%;两个加号“++”对应于45℃保温15min后,突变体蛋白的残余活性保留45.1~60.0%;三个加号“+++”对应于45℃保温15min后,突变体蛋白的残余活性保留60.1~80.0%。
表2:7β-羟基甾醇脱氢酶突变体序列和相应的活性改进列表
对应突变体编号的7β-羟基甾醇脱氢酶的氨基酸序列分别如下:
M10:将如序列表中SEQ ID No.2所示氨基酸序列的第57位甘氨酸替换为苏氨酸,第128位甘氨酸替换为甲硫氨酸,第176位丝氨酸替换为丙氨酸,第197位天冬酰胺替换为苏氨酸,第258位甘氨酸替换为苏氨酸;
M11:将如序列表中SEQ ID No.2所示氨基酸序列的第57位甘氨酸替换为苏氨酸,第128位甘氨酸替换为丙氨酸,第172位丙氨酸替换为异亮氨酸,第197位天冬酰胺替换为酪氨酸,第200位甘氨酸替换为天冬酰胺;
M12:将如序列表中SEQ ID No.2所示氨基酸序列的第163位异亮氨酸替换为缬氨酸,第172位丙氨酸替换为异亮氨酸,第197位天冬酰胺替换为酪氨酸,第243位天冬酰胺替换为色氨酸。
实施例3 重组E.coli BL21(DE3)/pET28a-7β-M6的表达
将实施例1中获得的突变体M6的重组大肠杆菌E.coli BL21(DE3)/pET28a-7β-M6,接种至含50μg/ml硫酸卡那霉素的LB培养基中,37℃振荡培养至OD600达到1.2,按1%(v/v)的接种量接入装有100ml LB培养基(含50μg/ml硫酸卡那霉素)的500ml三角烧瓶中,置于37℃、180rpm摇床振荡培养,当培养液的OD600达到0.6时,加入终浓度为0.2mmol/L的异丙基-β-D-硫代半乳糖苷(IPTG)作为诱导剂,16℃诱导24h。将培养液以8000×g离心10min,收集细胞,并用生理盐水洗涤两次。将100ml培养液中获得的细胞悬浮于10ml的磷酸钾缓冲液(100mM,pH 8.0)中,超声破碎,12000×g离心收集上清液,粗酶液的活性为51U/ml。另外,将收获的细胞冷冻干燥,获得的冻干细胞的活性为3.8U/mg DCW。
实施例4 重组P.pastoris/pPICZαA-7β-M6的构建和表达
依据实施例1获得的M6突变体的序列,提交金斯瑞生物科技有限公司对其进行密码子优化,使其适合于在毕赤酵母中进行分泌表达。优化后的核酸序列如序列表中SEQ ID No.5所示,对其进行序列全合成。设计上下游引物SEQ ID No.6和SEQ ID No.7,对合成的序列进行PCR扩增,用限制性内切酶EcoR I和Not I进行双酶切,将酶切片段与同样使用限制性内切酶EcoR I和Not I双酶切的pPICZαA质粒进行连接,获得重组质粒pPICZαA-7β-M6,然后在37℃,对其用限制性内切酶Sac I双酶切4h,使其线性化。
将80μl毕赤酵母X33的感受态细胞和1μg线性化质粒DNA样品混匀,转移至预冷的电转杯(电极间距0.2cm)中,冰浴5min,然后在2kV、5ms的条件下脉冲电击一次,向电转杯中迅速加入0.5ml冰上预冷的山梨醇溶液(1M),然后将电转杯中的菌液转移到装有0.5ml YPD液体培养基(蛋白胨20g/L,酵母提取物10g/L,葡萄糖20g/L,pH 6.0)的1.5ml Eppendorf管中,于30℃、200rpm培养2h;用移液枪吸取200μl电转化复苏后的菌液,涂布于YPDZ固体培养基平板(蛋白胨20g/L,酵母提取物10g/L,葡萄糖20g/L,博莱霉素1mg/ml,琼脂粉20g/L,pH 6.0),倒置于30℃培养箱中培养2天左右,至有肉眼可见的转化子长出,获得重组毕赤酵母P.pastoris X33/pPICZαA-7β-M6。
将重组毕赤酵母P.pastoris X33/pPICZαA-7β-M6接种至YPDZ液体培养基(蛋白胨20g/L,酵母提取物10g/L,葡萄糖20g/L,博莱霉素100μg/ml,pH 6.0)中,于30℃,250rpm震荡培养24h,按1%的接种量接种至100ml含有100μg/ml氨苄青霉素的BMGY液体培养基(蛋白胨20g/L,酵母提取物10g/L,甘油10g/L,无氨基酸酵母氮源13.6g/L,生物素0.4mg/L,终浓度为100mM的磷酸钾缓冲盐,pH 6.0)中,置于30℃,250rpm摇床中培养,当培养液的光密度OD600达到1.5时,停止培养,静置2h使酵母细胞沉降,小心倾倒出BMGY培养基上清,然后将收集的菌体用100ml的BMMY培养基(甲醇10ml/L,蛋白胨20g/L,酵母提取物10g/L,生物素0.4mg/L,无氨基酸酵母氮源13.6g/L,终浓度为100mM的磷酸钾缓冲盐,pH 6.0)重新悬浮,置于30℃、250rpm摇床中继续培养,每24h添加1ml的纯甲醇进行诱导,持续培养、诱导96h。培养结束后,将培养液于4℃、8000×g离心去除菌体,获得的上清粗酶液的活性为12U/ml。
实施例5 重组羟基甾醇脱氢酶7β-M6的固定化
取100ml如实施例4所述的重组羟基甾醇脱氢酶7β-M6的粗酶液,加入5g环氧树脂ES-103树脂,20℃、180rpm振摇20h,过滤,然后用100ml磷酸盐缓冲液(100mM,pH 5.5)洗涤3次,获得固定化酶7β-M6@ES-103,比活力为137U/g树脂,酶活上载率为57.3%。
实施例6 重组羟基甾醇脱氢酶7β-M6与葡萄糖脱氢酶的共固定化
取500U如实施例3所述的重组羟基甾醇脱氢酶7β-M6和500U葡萄糖脱氢酶(葡萄糖脱氢酶的重组表达参见:中国现代医学杂志,2007,17(2):172-174),用磷酸钾缓冲液(100mM,pH 5.5)稀释至100ml,加入5g环氧树脂ES-103树脂,20℃、180rpm振摇20h,过滤,然后用100ml磷酸盐缓冲液(100mM,pH5.5)洗涤3次,获得重组7β-M6和葡萄糖脱氢酶的共固定化酶7β-M6/GDH@ES-103,其中7β-M6的比活力为61U/g树脂,葡萄糖脱氢酶的比活力为69U/g树脂。
实施例7 重组7α-HSDHCa与乳酸脱氢酶的共固定化
取1000U重组7α-HSDHCa(7α-HSDHCa的重组表达参见:Appl Microbiol Biotechnol,2012,95,1221–1233)和1000U乳酸脱氢酶(乳酸脱氢酶的重组表达参见:FEBS Lett,1991,290,61–64)的粗酶液,用磷酸钾缓冲液(100mM,pH 5.5)稀释至100ml,加入5g环氧树脂ES-103树脂,20℃、180rpm振摇20h,过滤,然后用100ml磷酸盐缓冲液(100mM,pH 5.5)洗涤3次,获得重组7α-HSDHCa和乳酸脱氢酶的共固定化酶7α-HSDHCa/LDH@ES-103,其中7α-HSDHCa的比活力为135U/g树脂,乳酸脱氢酶的比活力为116U/g树脂。
实施例7,8 重组7β-羟基甾醇脱氢酶催化合成UDCA
将1.2g 7-KLCA溶解在10ml磷酸钾缓冲液(100mM,pH8.0)中,依次加入终浓度0.5mM的NADP+、20U的葡萄糖脱氢酶液、0.81g无水葡萄糖,然后分别加入20U的重组7β-M0(J Agric Food Chem,2017,65,1178-1185)或者20U如实施例4获得的重组羟基甾醇脱氢酶7β-M6。反应在35℃、180rpm振荡混匀条件下进行,间歇取样检测反应转化率,至转化率不再继续增长时停止反应,取样测定酶的残余活力。转化率检测使用C-18柱,甲醇/水=75:25(磷酸调pH=3)为流动相,柱温30℃,流速0.8ml/mi,检测波长210nm。
表3 重组7β-羟基甾醇脱氢酶催化7-KLCA转化合成UDCA
结果如表3,反应10h后,7β-M0对底物的转化率为98.0%,残余活力还剩余21%。而7β-M6在反应6个小时之后,转化率已经高于99.5%,残余活力为78%。结果表明,突变体7β-M6由于具有更高的活性和稳定性,催化合成UDCA的效果显著优于母本7β-M0。
实施例9 重组羟基甾醇脱氢酶7β-M6催化合成UDCA
在20ml夹套反应器中,加入10ml磷酸钾缓冲液(100mM,pH 8.0),加入105mg如实施例3制备的重组E.coli BL21(DE3)/pET28a-7β-M6冻干细胞,400U葡萄糖脱氢酶,终浓度0.05mM的NADP+,加入1.2g 7-KLCA,30℃条件下磁力搅拌反应。通过自动电位滴定仪控制滴加NaOH溶液(1.0M),维持反应液pH在8.0左右。间歇取样检测反应转化率,反应30min后,反应转化率99.8%。终止反应,离心分离除去细胞,用1mol/L的HCL调节pH至3~4使UDCA析出,用等体积的乙酸乙酯萃取三次,萃取液混合,用等体积饱和食盐水洗涤两次,无水硫酸钠干燥过夜,然后旋转蒸发除去溶剂,得到1.14g白色固体,纯度高于95%。
实施例10 重组羟基甾醇脱氢酶7β-M6催化合成UDCA
在2L三颈烧瓶中,加入1L磷酸钾缓冲液(100mM,pH 8.0),0.26g如实施例3制备的重组E.coli BL21(DE3)/pET28a-7β-M6冻干细胞,1000U葡萄糖脱氢酶,终浓度0.1mM的NADP+,以及40g 7-KLCA,30℃条件下机械搅拌反应,搅拌转速为200rpm。通过自动电位滴定仪控制滴加1.0N的NaOH溶液,维持反应液pH在8.0左右。间歇取样检测反应转化率,反应5h后,反应转化率高于99.9%,终止反应,用1mol/L的HCL调节pH至3~4使UDCA析出,用等体积的乙酸乙酯萃取三次,将萃取液混合,用等体积饱和食盐水洗涤两次,洗涤后的萃取液用无水硫酸钠干燥过夜,然后旋转蒸发浓缩至有结晶析出,冷却至室温,抽滤除去残余溶剂,烘干至恒重,得到37.6g白色固体,纯度高于99%。
实施例11 固定化酶7β-M6@ES-103催化合成UDCA
将1.2g 7-KLCA溶解在10ml磷酸钾缓冲液(100mM,pH8.0)中,依次加入终浓度0.5mM的NADP+、20U的葡萄糖脱氢酶、0.81g无水葡萄糖,然后加入0.2g如实施例5制备的固定化酶7β-M6@ES-103。反应在30℃,180rpm振荡混匀条件下进行,间歇取样检测反应转化率。转化6h,转化率高于99.5%。
实施例12 共固定化酶7β-M6/GDH@ES-103催化合成UDCA
将1.2g 7-KLCA溶解在10ml磷酸钾缓冲液(100mM,pH8.0)中,依次加入终浓度0.5mM的NADP+、0.81g无水葡萄糖,然后加入1g如实施例6制备的共固定化酶7β-M6/GDH@ES-103。反应在30℃,180rpm振荡混匀条件下进行,间歇取样检测反应转化率。反应3h,转化率已经高于99.5%。
实施例13 共固定化酶耦联催化合成UDCA
称取5g如实施例6制备的共固定化酶7β-M6/GDH@ES-103和3g如实施例7制备的共固定化酶7α-HSDH/LDH@ES-103,分别装填到带夹套和筛板的填充柱中,夹套中通恒温水,控温30℃。
将4g CDCA溶解在100ml磷酸钾缓冲液(100mM,pH8.0)中,依次加入终浓度为0.5mM的NAD+和NADP+、0.27g无水葡萄糖和0.14g丙酮酸。通过蠕动泵使底物溶液依次流过7α-HSDH/LDH@ES-103和7β-HSDH-M6/GDH@ES-103的填充柱,溶液流速1.0ml/min,检测流出液的反应转化率,高于99.5%。
收集流出的反应液,用1mol/L的HCL调节pH至3~4使UDCA析出,用等体积的乙酸乙酯萃取三次,将三次萃取液混合,用等体积饱和食盐水洗涤两次,洗涤后的萃取液用无水硫酸钠干燥过夜,然后旋转蒸发除去溶剂,得到3.8g白色固体,纯度高于99%。
上述的对实施例的描述是为便于该技术领域的普通技术人员能理解和使用发明。熟悉本领域技术的人员显然可以容易地对这些实施例做出各种修改,并把在此说明的一般原理应用到其他实施例中而不必经过创造性的劳动。因此,本发明不限于上述实施例,本领域技术人员根据本发明的揭示,不脱离本发明范畴所做出的改进和修改都应该在本发明的保护范围之内。
<110> 华东理工大学、苏州百福安酶技术有限公司
<120> 7β-羟基甾醇脱氢酶突变体及其在熊脱氧胆酸合成中的应用
<160> 7
<170> PatentIn version 3.5
<210> 1
<211> 795
<212> DNA
<213> 扭链瘤胃球菌 (Ruminococcus torques)
<400> 1
atgaatctgc gtgaaaaata cggcgaatgg ggcatcattc tgggcgcgac cgagggcgtg 60
ggcaaggcgt ttgcggaaaa gattgcgagc gaaggcatga gcgtggtcct ggtgggccgt 120
cgtgaagaaa aactgcaaga actgggtaaa tctattagcg aaacctatgg cgtggatcat 180
atggtcattc gtgccgattt cgcgcaaagc gattgcaccg acaagatctt tgaagcgacc 240
aaagatctgg acatgggctt tatgagctat gtggcgtgtt ttcacacctt tggcaagctg 300
caggataccc cgtgggaaaa acatgaacag atgattaatg tgaacgtgat gacctttctg 360
aagtgttttt accattatat gggcatcttt gcgaaacagg atcgtggcgc ggtgatcaat 420
gtgagcagcc tgaccgcgat tagcagcagc ccgtataatg cgcagtatgg cgcaggcaag 480
agctacatca aaaagctgac cgaagcggtg gcagcggaat gcgaaagcac caatgtggat 540
gtggaagtga ttaccctggg caccgtcatt accccgagcc tgctgagcaa tctgccaggt 600
ggcccagcag gtgaagcaat gatgaaaacg gcgatgaccc cggaagcgtg cgtggaagaa 660
gcgtttgata atctgggcaa aagcctgagc gttattgcgg gcgaacataa caaagccaat 720
gttcataatt ggcaggcgaa caaaaccgat gatgaatata tccgttacat gggtagcttc 780
tatagcaaca attaa795
<210> 2
<211> 264
<212> PRT
<213> 扭链瘤胃球菌 (Ruminococcus torques)
<400> 2
Met Asn Leu Arg Glu Lys Tyr Gly Glu Trp Gly Ile Ile Leu Gly
1 5 1015
Ala Thr Glu Gly Val Gly Lys Ala Phe Ala Glu Lys Ile Ala Ser
202530
Glu Gly Met Ser Val Val Leu Val Gly Arg Arg Glu Glu Lys Leu
354045
Gln Glu Leu Gly Lys Ser Ile Ser Glu Thr Tyr Gly Val Asp His
505560
Met Val Ile Arg Ala Asp Phe Ala Gln Ser Asp Cys Thr Asp Lys
657075
Ile Phe Glu Ala Thr Lys Asp Leu Asp Met Gly Phe Met Ser Tyr
808590
Val Ala Cys Phe His Thr Phe Gly Lys Leu Gln Asp Thr Pro Trp
95100 105
Glu Lys His Glu Gln Met Ile Asn Val Asn Val Met Thr Phe Leu
110 115 120
Lys Cys Phe Tyr His Tyr Met Gly Ile Phe Ala Lys Gln Asp Arg
125 130 135
Gly Ala Val Ile Asn Val Ser Ser Leu Thr Ala Ile Ser Ser Ser
140 145 150
Pro Tyr Asn Ala Gln Tyr Gly Ala Gly Lys Ser Tyr Ile Lys Lys
155 160 165
Leu Thr Glu Ala Val Ala Ala Glu Cys Glu Ser Thr Asn Val Asp
170 175 180
Val Glu Val Ile Thr Leu Gly Thr Val Ile Thr Pro Ser Leu Leu
185 190 195
Ser Asn Leu Pro Gly Gly Pro Ala Gly Glu Ala Met Met Lys Thr
200 205 210
Ala Met Thr Pro Glu Ala Cys Val Glu Glu Ala Phe Asp Asn Leu
215 220 225
Gly Lys Ser Leu Ser Val Ile Ala Gly Glu His Asn Lys Ala Asn
230 235 240
Val His Asn Trp Gln Ala Asn Lys Thr Asp Asp Glu Tyr Ile Arg
245 250 255
Tyr Met Gly Ser Phe Tyr Ser Asn Asn
260
<210> 3
<211> 30
<212> DNA
<213> 人工序列
<400> 3
ccggaattca tgaatctgcg tgaaaaatac30
<210> 4
<211> 29
<212> DNA
<213> 人工序列
<400> 4
ccgctcgagt taattgttgc tatagaagc 29
<210> 5
<211> 792
<212> DNA
<213> 人工序列
<400> 5
atgaatttga gagaaaagta cggagagtgg ggtattattt tgggtgctac tgaaggtgtt 60
ggtaaagctt tcgctgaaaa gattgcttct gagggaatgt ctgttgtttt ggttggtaga 120
agagaagaga agttgcaaga attgggtaaa tctatttctg agacttacac tgttgatcat 180
atggttatta gagctgattt tgctcaatct gattgtactg ataagatctt cgaagctact 240
aaggatttgg atatgggttt tatgtcttac gttgcttgtt tccatacttt cggtaaattg 300
caagatactc catgggaaaa acacgagcaa atgatcaacg ttaacgttat gactttcttg 360
aagtgtttct accactacat gggtatcttc gctaagcaag atagaggtgc tgttattaat 420
gtttcttctt tgactgctat ctcttcttct ccttacaacg ctcaatatgg tgctggtaaa 480
tcttacatta agaaattgac tgaagctgtt gctttggagt gtgagtctac taacgttgat 540
gttgaggtta ttactttggg tactgttatt actccatctt tgttgtctaa cttgccaggt 600
ggtcctgctg gtgaagctat gatgaagact gctatgactc ctgaggcttg tgttgaagag 660
gctttcgata atttgggtaa atctttgtct gttattgctg gtgaacataa caaggctaat 720
gttcacaact ggcaagctaa caaaactgat gatgagtaca tcagatatat gggttctttt 780
tattctaaca at 792
<210> 6
<211> 34
<212> DNA
<213> 人工序列
<400> 6
gaattcatga atttgagaga aaagtacgga gagt 34
<210> 7
<211> 44
<212> DNA
<213> 人工序列
<400> 7
aaggaaaaaa gcggccgcat tgttagaata aaaagaaccc atat44
7β-羟基甾醇脱氢酶突变体及其在熊脱氧胆酸合成中的应用专利购买费用说明
![]()
Q:办理专利转让的流程及所需资料
A:专利权人变更需要办理著录项目变更手续,有代理机构的,变更手续应当由代理机构办理。
1:专利变更应当使用专利局统一制作的“著录项目变更申报书”提出。
2:按规定缴纳著录项目变更手续费。
3:同时提交相关证明文件原件。
4:专利权转移的,变更后的专利权人委托新专利代理机构的,应当提交变更后的全体专利申请人签字或者盖章的委托书。
Q:专利著录项目变更费用如何缴交
A:(1)直接到国家知识产权局受理大厅收费窗口缴纳,(2)通过代办处缴纳,(3)通过邮局或者银行汇款,更多缴纳方式
Q:专利转让变更,多久能出结果
A:著录项目变更请求书递交后,一般1-2个月左右就会收到通知,国家知识产权局会下达《转让手续合格通知书》。

















动态评分
0.0