









专利摘要
本发明属于基因工程领域,具体涉及一种3‑甾酮‑△1‑脱氢酶及其编码基因和应用。本发明研究首次发现一种全新的3‑甾酮‑△1‑脱氢酶,其在转化培养基中底物4‑AD浓度高达0.8%时,转化率可高达99%,转化液中产物ADD的浓度达到7.9g/L,远远高于其他相关报道,有效的减少底物4‑AD残留,提高产物ADD的含量,减少了分离提取的步骤,具有很好的产业化前景。
权利要求
1.一种分离的3-甾酮-△
2.一种分离的多核苷酸,所述分离的多核苷酸编码如权利要求1所述的3-甾酮-△
3.如权利要求2所述的分离的多核苷酸,其特征在于,所述多核苷酸的核苷酸序列如SEQ ID NO:1所示。
4.一种重组表达载体,包含如权利要求2或3所述的分离的多核苷酸。
5.如权利要求4所述的重组表达载体,其特征在于,所述重组表达载体由如权利要求2或3所述的分离的多核苷酸克隆到表达载体中获得。
6.如权利要求5所述的重组表达载体,其特征在于,所述表达载体为pET-28a。
7.一种宿主细胞,所述细胞含有如权利要求4~6之任一项所述的重组表达载体或基因组中整合有外源的如权利要求2~3之任一项所述的分离的多核苷酸。
8.一种制备如权利要求1所述的3-甾酮-△
在合适的条件下培养如权利要求7所述的宿主细胞,使之表达所述3-甾酮-△
9.如权利要求1所述3-甾酮-△
10.一种4-AD生成ADD的转化剂,含有如权利要求1所述3-甾酮-△
11.一种转化4-AD生成ADD的方法,包括步骤:将如权利要求1所述的3-甾酮-△
说明书
技术领域
本发明属于基因工程领域,具体涉及一种3-甾酮-△
背景技术
甾体化合物(steroids)又称类固醇,是广泛存在于生物体组织内的一类重要的天然有机化合物。从结构上看,甾体是一类以环戊烷多氢菲(Cl7)为母核的化合物,由三个六元环和一个五元环组成,分别称为A、B、C、D环(如图1所示)。由于母核上取代基、双键位置或立体构型的不同,形成了种类繁多的甾体化合物,对机体起着非常重要的调节作用。甾体类药物是指具有甾核结构的甾体激素类药物,包括盐皮质激素、糖皮质激素、性激素和蛋白同化激素等。盐皮质激素和糖皮质激素具有抗炎症、抗过敏等功效,广泛用于治疗类风湿性关节炎、支气管哮喘和湿疹等疾病;性激素是治疗雄性器官衰退和一些妇科疾病的主要药物,是治疗乳腺癌、前列腺癌的辅助治疗药物,也是近年来需求旺盛的口服避孕药的主要成分;蛋白同化激素可促进蛋白质合成并抑制蛋白质异化,使肌肉生长;促使韩、碟在组织中沉积,加速骨组织分化和生长等。由于甾体类药物对机体起着非常重要的调节作用,到目前为止,已获得批准使用的甾体类药物大约有300多种,是产量仅次于抗生素的第二大类药物。
3-甾酮-△
综上所述,目前利用基因工程方法将不同来源的KSDD酶基因外源表达到不同宿主中,构建重组菌株进行生物转化4-AD制备ADD仍然面临底物投料浓度小、转化率低等问题。因此充分挖掘KSDD酶基因资源,构建高效表达的KSDD酶基因工程菌株对实现利用生物技术转化AD制备重要甾体类原料药ADD的工业化生产具有重要意义。
发明内容
为了克服现有技术中所存在的问题,本发明的目的在于提供一种新的3-甾酮-△
为了实现上述目的以及其他相关目的,本发明采用如下技术方案:
本发明的第一方面,提供一种分离的3-甾酮-△
如本发明实施例中所列举的,所述3-甾酮-△
本发明的第二方面,提供一种分离的多核苷酸,所述分离的多核苷酸编码前述3-甾酮 -△
本发明的编码前述3-甾酮-△
本发明的编码所述3-甾酮-△
如本发明实施例中所列举的,所述编码前述3-甾酮-△
本发明的第三方面,提供一种重组表达载体,包含前述分离的多核苷酸。
本领域的技术人员熟知的方法能用于构建所述重组表达载体。这些方法包括重组DNA 技术、DNA合成技术等。可将编码所述3-甾酮-△
本发明的第四方面,提供一种宿主细胞,所述细胞含有前述重组表达载体或基因组中整合有外源的前述分离的多核苷酸。
本发明的较佳案例中,所述宿主细胞采用大肠杆菌,枯草芽孢杆菌或毕赤酵母。所述的大肠杆菌优选为BL21(DE3)。所述枯草芽孢杆菌优选为BS158。所述毕赤酵母优选为GS115。
本发明的第五方面,提供一种制备前述3-甾酮-△1-脱氢酶的方法,包括以下之任一:
(1)构建含有3-甾酮-△
或者
(2)在合适的条件下培养前述宿主细胞,使之表达所述3-甾酮-△
本发明的较佳案例中,所述表达载体采用pET-28a。所述宿主细胞采用大肠杆菌。所述的大肠杆菌优选为BL21(DE3)。
本发明的第六方面,提供第一方面所述3-甾酮-△
本发明的第七方面,提供一种4-AD生成ADD的转化剂,含有前述第一方面所述3-甾酮-△
需要说明的是,在本发明中,4-AD均为雄烯二酮,ADD均为雄二烯二酮。
本发明的第八方面,提供一种转化4-AD生成ADD的方法,包括步骤:将本发明的第一方面的3-甾酮-△
与现有技术相比,本发明具有如下有益效果:
1、发明人首次从新金分支杆菌DSM 1381中克隆出一个新的3-甾酮-△
2、通过基因工程手段,将kstD2基因在大肠杆菌中进行了异源过量表达,获得了高效表达3-甾酮-△
附图说明
图1:甾体化合物通式,在本发明中,3-甾酮甾体化合物可以是现有3-甾酮-4-烯甾体类化合物,常见的有AD、9α-OH-AD、17α-羟基黄体酮、环氧黄体酮、霉菌氧化物、可的松、氢化可的松、依普利酮、20-(羟甲基)孕-4-烯-3-酮。
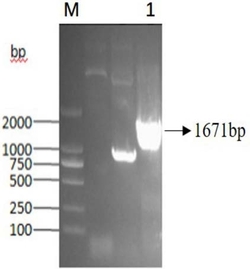
图2:琼脂糖凝胶电泳检测KstD2 PCR产物。
图3:重组大肠杆菌BL21-kstD2 AD转化率。
图4:重组大肠杆菌BL21-kstD2AD转化实验中的AD的剩余量。
图5:重组大肠杆菌BL21-kstD2 AD转化实验中ADD的产量。
具体实施方式
在进一步描述本发明具体实施方式之前,应理解,本发明的保护范围不局限于下述特定的具体实施方案;还应当理解,本发明实施例中使用的术语是为了描述特定的具体实施方案,而不是为了限制本发明的保护范围。下列实施例中未注明具体条件的试验方法,通常按照常规条件,或者按照各制造商所建议的条件。
当实施例给出数值范围时,应理解,除非本发明另有说明,每个数值范围的两个端点以及两个端点之间任何一个数值均可选用。除非另外定义,本发明中使用的所有技术和科学术语与本技术领域技术人员通常理解的意义相同。除实施例中使用的具体方法、设备、材料外,根据本技术领域的技术人员对现有技术的掌握及本发明的记载,还可以使用与本发明实施例中所述的方法、设备、材料相似或等同的现有技术的任何方法、设备和材料来实现本发明。
除非另外说明,本发明中所公开的实验方法、检测方法、制备方法均采用本技术领域常规的分子生物学、生物化学、染色质结构和分析、分析化学、细胞培养、重组DNA技术及相关领域的常规技术。这些技术在现有文献中已有完善说明,具体可参见Sambrook等MOLECULAR CLONING:A LABORATORY MANUAL,Second edition,Cold Spring HarborLaboratory Press,1989 and Third edition,2001;Ausubel等,CURRENT PROTOCOLS INMOLECULAR BIOLOGY,John Wiley&Sons,New York,1987 and periodic updates;theseries METHODS IN ENZYMOLOGY,Academic Press,San Diego;Wolffe,CHROMATINSTRUCTURE AND FUNCTION,Third edition,Academic Press,San Diego,1998;METHODSIN ENZYMOLOGY,Vol.304,Chromatin(P.M.Wassarman and A.P.Wolffe,eds.),AcademicPress,San Diego,1999;和METHODS IN MOLECULAR BIOLOGY,Vol.119,ChromatinProtocols(P.B.Becker,ed.)Humana Press,Totowa,1999等。
实施例1新金分支杆菌DSM 1381 3-甾酮-△
利用Illumina Miseq测序技术对新金分支杆菌DSM 1381进行测序分析,根据基因注释信息,得到完整的3-甾酮-△
GTGACCGATCAGAACAACATCACCGTCGACCTCGTCGTCGTCGGCTCGGGTACCGGGATGGCGGCAGCATTGGCTGCCCACGAGCTGGGAATGTCGACGCTGATCGTCGAGAAGAGCGCCTATGTCGGTGGTTCGACGGCTCGCTCCGGCGGTGCCTTCTGGCTTCCCGGCAGCTCCATTCTCAAGGACGCCGGTTCGGCGGACACTCCGGCCAAGGCGCGCACCTACCTTGAAGCACTCGTCGGTGACGACGTCTCACCCGAACGCGCACGCACTTTCATCGATCAGATCCCCGCGACCATCGACATGTTGCGTCGCACCACCCCGATGAAGTTCATGTGGGCCAAGGGATATTCGGACTACCACCCGGAGAGGCCAGGAGGCAGTGCGGTGGGCCGGACCTGTGAGTGTCGCCCGTTCGACACTGCGGTCCTCGGTCCAGAGCTGGCGCGGCTACGACCTGGAGTGATGAAGTCATCGTTCCCGATGCCGGTCACCGGCGCCGATTACCGTTGGCTGAACCTGATGGCCCGCACCCCGCGCAAGTCCTGGCCGCGGATCATGCTGCGGGCCATGCAGGGTGTCGGCGGTTTGGCCCTGCGGCGCCGGTACGCCGCAGGCGGCCAGGCCTTGGCGGCCGGGATGTTCGCCGGCGTGCTGCAGGCGGGGATCCCGGTGTGGACCGATTCGACGGTGACCGAGCTCATCACCGATGGTGGGCGGGTGACCGGCGCGCGGGTGCTGCGCGAGGGATCGGCCGTGACCGTCACCGCACGCCGTGGCATCGTGCTGGCCACCGGCGGTTTCGACCACGAGATGAATTGGCGGCGGAAGTTCCAGTCCGAGCTCCTCGGTGAACATCTCAGCCTTGGGGCCGAGAGCAATACCGGCGATGGCATCCGGCTCGCCCAGGACCTGGGCGCAGGCACCGGACTGATGGACCAGGCATGGTGGTTTCCGGCCTTTGCTCCGCTGCCTGGCGGGGATCCCACCGTGATGCTGGCCGAGCGGTCGCTGCCCGGCTGCCTGCTGGTAGACCAGACCGGTGAGCGCTTCATCAACGAGGCCACCGACTACATGTCCTTCGGACAGCAGCTGCTGCGTCGCGAACACGCGGGCAATCCGGTCGAGACGATGTGGATGATCTTCGATCAGCGCTACCGGAACAGCTATCTGCTTGCCGCCGAACTATTTCCACGAATGCCGATCCCACAGAGTTGGTACGACGCCGGGATCGCGCACCGCGGCACGGATGCGGAAGCACTGGGCCGCCAGATCGGTTTCGATCCCGCGACGTTGGTCGCCACGATCGAGCGGTTCAACGGACTCGCCGATGCCGGTGTCGACGCCGACTTCCAGCGCGGCGCGAGCGCCTACGACCGCTACTACGGCGACCCGACGATCACGCCCAACCCGAACCTGCGACCGCTGGATCCCGGCCCGCTGTACGCCGTCAAGGTCGTGCTGAGCGACCTGGGCACCTGTGGTGGGGTCCTGTGCGACGTGAACGGCCGGGTTCTGCGCGAAGACGGAGTGCCCATCGACGGTCTGTACGCGATCGGCAATACCGCGGCCAACGCATTCGGCAAGACCTACCCGGGCGCGGGCGCGACCATCGCGCAGGGGCTGGTGTACGGCCATGTTGCCGCGCAGCATGCCGC CGGACACACCTGA。
所述3-甾酮-△
VTDQNNITVDLVVVGSGTGMAAALAAHELGMSTLIVEKSAYVGGSTARSGGAFWLPGSSILKDAGSADTPAKARTYLEALVGDDVSPERARTFIDQIPATIDMLRRTTPMKFMWAKGYSDYHPERPGGSAVGRTCECRPFDTAVLGPELARLRPGVMKSSFPMPVTGADYRWLNLMARTPRKSWPRIMLRAMQGVGGLALRRRYAAGGQALAAGMFAGVLQAGIPVWTDSTVTELITDGGRVTGARVLREGSAVTVTARRGIVLATGGFDHEMNWRRKFQSELLGEHLSLGAESNTGDGIRLAQDLGAGTGLMDQAWWFPAFAPLPGGDPTVMLAERSLPGCLLVDQTGERFINEATDYMSFGQQLLRREHAGNPVETMWMIFDQRYRNSYLLAAELFPRMPIPQSWYDAGIAHRGTDAEALGRQIGFDPATLVATIERFNGLADAGVDADFQRGASAYDRYYGDPTITPNPNLRPLDPGPLYAVKVVLSDLGTCGGVLCDVNGRVLREDGVPIDGLYAIGNTAANAFGKTYPGAGATIAQGLVYGHVAAQHAAGHT。
与目前报道的其它3-甾酮-△
kstD2-F 5’cagc
kstD2-R 5’ctcg
其中,在引物kstD2-F上引入EcoRI酶切位点,在引物kstD2-R上引入Hind III酶切位点。
以新金分支杆菌DSM 1381的总DNA为模板,以引物kstD2-F和kstD2-R为扩增引物,选择高保真PrimeSTAR HS DNA Polymerase with GC Buffer(TaKaRa)进行PCR扩增具体条件如下:
95℃预变性3min,95℃变性30sec,62℃退火30sec,72℃延伸2min,30个循环;72℃最后延伸10min;用1.5%琼脂糖凝胶电泳检测PCR产物,结果如图2所示。目标DNA片段大小为1674bp,与目标基因片段长度一直。经测序公司测序分析,得到3-甾酮-△
实施例2 3-甾酮-△
将实施例1得到的片段经EcoRI和Hind III双酶切,与同样利用EcoRI和Hind III进行双酶切的表达载体pET-28a进行连接,连接产物经转化大肠杆菌DH5α感受态细胞(购自TAKARA公司),筛选验证得到表达重组质粒。将重组质粒送到苏州金唯智生物科技有限公司进行测序分析,经测序所述3-甾酮-△
实施例3基因工程菌的酶活力测定
用IPTG诱导24h后,6000rpm,4℃离心10分钟从50Ml培养物中收集重组大肠杆菌BL21(DE)和枯草芽孢杆菌164菌体,然后用50mM Tris-HCl缓冲区(pH值7.0)两次清洗,最后用4mL缓冲液重悬。重悬物进行超声波破碎处理,然后12000rpm 4℃离心30分钟。取上清测定KstD酶活。酶活测定使用分光光度法,利用NanoDrop 2000测定600nm(ε600 nm=18.7×103cm
实施例4基因工程菌株BL21-kstD2对AD的转化
将重组大肠杆菌BL21-kstD2以5%接种量接种于TB培养基中,在37℃200rpm摇床中培养,当OD600达到0.8左右时,加入异丙基硫代半乳糖昔(IPTG)使其终浓度达到1.0 mM,同时按照1.0%(W/V)加入4-AD和0.5%(W/V)加入羟丙基环糊精,继续在37℃转化。经HPLC检测分析,转化21h后底物AD的转化率达到99%(见图3),重组大肠杆菌BL21-kstD2AD转化实验中的AD的剩余量如图4所示,产物ADD含量达到7.9g/L(见图5)。
以上所述,仅为本发明的较佳实施例,并非对本发明任何形式上和实质上的限制,应当指出,对于本技术领域的普通技术人员,在不脱离本发明方法的前提下,还将可以做出若干改进和补充,这些改进和补充也应视为本发明的保护范围。凡熟悉本专业的技术人员,在不脱离本发明的精神和范围的情况下,当可利用以上所揭示的技术内容而做出的些许更动、修饰与演变的等同变化,均为本发明的等效实施例;同时,凡依据本发明的实质技术对上述实施例所作的任何等同变化的更动、修饰与演变,均仍属于本发明的技术方案的范围内。
序列表
<110> 中国科学院上海高等研究院
<120> 一种3-甾酮-△1-脱氢酶及其编码基因和应用
<130> 181606
<160> 4
<170> SIPOSequenceListing 1.0
<210> 1
<211> 1674
<212> DNA
<213> 人工序列(Artificial Sequence)
<400> 1
gtgaccgatc agaacaacat caccgtcgac ctcgtcgtcg tcggctcggg taccgggatg 60
gcggcagcat tggctgccca cgagctggga atgtcgacgc tgatcgtcga gaagagcgcc 120
tatgtcggtg gttcgacggc tcgctccggc ggtgccttct ggcttcccgg cagctccatt 180
ctcaaggacg ccggttcggc ggacactccg gccaaggcgc gcacctacct tgaagcactc 240
gtcggtgacg acgtctcacc cgaacgcgca cgcactttca tcgatcagat ccccgcgacc 300
atcgacatgt tgcgtcgcac caccccgatg aagttcatgt gggccaaggg atattcggac 360
taccacccgg agaggccagg aggcagtgcg gtgggccgga cctgtgagtg tcgcccgttc 420
gacactgcgg tcctcggtcc agagctggcg cggctacgac ctggagtgat gaagtcatcg 480
ttcccgatgc cggtcaccgg cgccgattac cgttggctga acctgatggc ccgcaccccg 540
cgcaagtcct ggccgcggat catgctgcgg gccatgcagg gtgtcggcgg tttggccctg 600
cggcgccggt acgccgcagg cggccaggcc ttggcggccg ggatgttcgc cggcgtgctg 660
caggcgggga tcccggtgtg gaccgattcg acggtgaccg agctcatcac cgatggtggg 720
cgggtgaccg gcgcgcgggt gctgcgcgag ggatcggccg tgaccgtcac cgcacgccgt 780
ggcatcgtgc tggccaccgg cggtttcgac cacgagatga attggcggcg gaagttccag 840
tccgagctcc tcggtgaaca tctcagcctt ggggccgaga gcaataccgg cgatggcatc 900
cggctcgccc aggacctggg cgcaggcacc ggactgatgg accaggcatg gtggtttccg 960
gcctttgctc cgctgcctgg cggggatccc accgtgatgc tggccgagcg gtcgctgccc 1020
ggctgcctgc tggtagacca gaccggtgag cgcttcatca acgaggccac cgactacatg 1080
tccttcggac agcagctgct gcgtcgcgaa cacgcgggca atccggtcga gacgatgtgg 1140
atgatcttcg atcagcgcta ccggaacagc tatctgcttg ccgccgaact atttccacga 1200
atgccgatcc cacagagttg gtacgacgcc gggatcgcgc accgcggcac ggatgcggaa 1260
gcactgggcc gccagatcgg tttcgatccc gcgacgttgg tcgccacgat cgagcggttc 1320
aacggactcg ccgatgccgg tgtcgacgcc gacttccagc gcggcgcgag cgcctacgac 1380
cgctactacg gcgacccgac gatcacgccc aacccgaacc tgcgaccgct ggatcccggc 1440
ccgctgtacg ccgtcaaggt cgtgctgagc gacctgggca cctgtggtgg ggtcctgtgc 1500
gacgtgaacg gccgggttct gcgcgaagac ggagtgccca tcgacggtct gtacgcgatc 1560
ggcaataccg cggccaacgc attcggcaag acctacccgg gcgcgggcgc gaccatcgcg 1620
caggggctgg tgtacggcca tgttgccgcg cagcatgccg ccggacacac ctga 1674
<210> 2
<211> 557
<212> PRT
<213> 人工序列(Artificial Sequence)
<400> 2
Val Thr Asp Gln Asn Asn Ile Thr Val Asp Leu Val Val Val Gly Ser
1 5 1015
Gly Thr Gly Met Ala Ala Ala Leu Ala Ala His Glu Leu Gly Met Ser
202530
Thr Leu Ile Val Glu Lys Ser Ala Tyr Val Gly Gly Ser Thr Ala Arg
354045
Ser Gly Gly Ala Phe Trp Leu Pro Gly Ser Ser Ile Leu Lys Asp Ala
505560
Gly Ser Ala Asp Thr Pro Ala Lys Ala Arg Thr Tyr Leu Glu Ala Leu
65707580
Val Gly Asp Asp Val Ser Pro Glu Arg Ala Arg Thr Phe Ile Asp Gln
859095
Ile Pro Ala Thr Ile Asp Met Leu Arg Arg Thr Thr Pro Met Lys Phe
100 105 110
Met Trp Ala Lys Gly Tyr Ser Asp Tyr His Pro Glu Arg Pro Gly Gly
115 120 125
Ser Ala Val Gly Arg Thr Cys Glu Cys Arg Pro Phe Asp Thr Ala Val
130 135 140
Leu Gly Pro Glu Leu Ala Arg Leu Arg Pro Gly Val Met Lys Ser Ser
145 150 155 160
Phe Pro Met Pro Val Thr Gly Ala Asp Tyr Arg Trp Leu Asn Leu Met
165 170 175
Ala Arg Thr Pro Arg Lys Ser Trp Pro Arg Ile Met Leu Arg Ala Met
180 185 190
Gln Gly Val Gly Gly Leu Ala Leu Arg Arg Arg Tyr Ala Ala Gly Gly
195 200 205
Gln Ala Leu Ala Ala Gly Met Phe Ala Gly Val Leu Gln Ala Gly Ile
210 215 220
Pro Val Trp Thr Asp Ser Thr Val Thr Glu Leu Ile Thr Asp Gly Gly
225 230 235 240
Arg Val Thr Gly Ala Arg Val Leu Arg Glu Gly Ser Ala Val Thr Val
245 250 255
Thr Ala Arg Arg Gly Ile Val Leu Ala Thr Gly Gly Phe Asp His Glu
260 265 270
Met Asn Trp Arg Arg Lys Phe Gln Ser Glu Leu Leu Gly Glu His Leu
275 280 285
Ser Leu Gly Ala Glu Ser Asn Thr Gly Asp Gly Ile Arg Leu Ala Gln
290 295 300
Asp Leu Gly Ala Gly Thr Gly Leu Met Asp Gln Ala Trp Trp Phe Pro
305 310 315 320
Ala Phe Ala Pro Leu Pro Gly Gly Asp Pro Thr Val Met Leu Ala Glu
325 330 335
Arg Ser Leu Pro Gly Cys Leu Leu Val Asp Gln Thr Gly Glu Arg Phe
340 345 350
Ile Asn Glu Ala Thr Asp Tyr Met Ser Phe Gly Gln Gln Leu Leu Arg
355 360 365
Arg Glu His Ala Gly Asn Pro Val Glu Thr Met Trp Met Ile Phe Asp
370 375 380
Gln Arg Tyr Arg Asn Ser Tyr Leu Leu Ala Ala Glu Leu Phe Pro Arg
385 390 395 400
Met Pro Ile Pro Gln Ser Trp Tyr Asp Ala Gly Ile Ala His Arg Gly
405 410 415
Thr Asp Ala Glu Ala Leu Gly Arg Gln Ile Gly Phe Asp Pro Ala Thr
420 425 430
Leu Val Ala Thr Ile Glu Arg Phe Asn Gly Leu Ala Asp Ala Gly Val
435 440 445
Asp Ala Asp Phe Gln Arg Gly Ala Ser Ala Tyr Asp Arg Tyr Tyr Gly
450 455 460
Asp Pro Thr Ile Thr Pro Asn Pro Asn Leu Arg Pro Leu Asp Pro Gly
465 470 475 480
Pro Leu Tyr Ala Val Lys Val Val Leu Ser Asp Leu Gly Thr Cys Gly
485 490 495
Gly Val Leu Cys Asp Val Asn Gly Arg Val Leu Arg Glu Asp Gly Val
500 505 510
Pro Ile Asp Gly Leu Tyr Ala Ile Gly Asn Thr Ala Ala Asn Ala Phe
515 520 525
Gly Lys Thr Tyr Pro Gly Ala Gly Ala Thr Ile Ala Gln Gly Leu Val
530 535 540
Tyr Gly His Val Ala Ala Gln His Ala Ala Gly His Thr
545 550 555
<210> 3
<211> 34
<212> DNA
<213> 人工序列(Artificial Sequence)
<400> 3
cagcaagctt gtgaccgatc agaacaacat cacc 34
<210> 4
<211> 28
<212> DNA
<213> 人工序列(Artificial Sequence)
<400> 4
ctcgaagctt tcaggtgtgt ccggcggc28
一种3-甾酮-△-脱氢酶及其编码基因和应用专利购买费用说明
![]()
Q:办理专利转让的流程及所需资料
A:专利权人变更需要办理著录项目变更手续,有代理机构的,变更手续应当由代理机构办理。
1:专利变更应当使用专利局统一制作的“著录项目变更申报书”提出。
2:按规定缴纳著录项目变更手续费。
3:同时提交相关证明文件原件。
4:专利权转移的,变更后的专利权人委托新专利代理机构的,应当提交变更后的全体专利申请人签字或者盖章的委托书。
Q:专利著录项目变更费用如何缴交
A:(1)直接到国家知识产权局受理大厅收费窗口缴纳,(2)通过代办处缴纳,(3)通过邮局或者银行汇款,更多缴纳方式
Q:专利转让变更,多久能出结果
A:著录项目变更请求书递交后,一般1-2个月左右就会收到通知,国家知识产权局会下达《转让手续合格通知书》。














动态评分
0.0