

IPC分类号 : C12N9/18,C12N15/63,C12P7/40,C12P35/00,C12P35/06





专利摘要
本发明公开了一种新型乙酰木聚糖酯酶的DNA,它的核苷酸序列如SEQAXENO.1所示。它的氨基酸序列如SEQAXENO.2所示。蛋白结构为同源三聚体时活性最高。本发明还公开了含上述新型乙酰木聚糖酯酶DNA的克隆载体和克隆技术,及重组酯酶对短链脂肪酸脂及酰基糖类的水解应用及其在生产β-内酰胺类抗生素的重要中间体-去酰基化的7-氨基头孢烷酸及去酰基化头孢菌素C中的应用,具有重要的工业应用价值。CCTCCNO:M20124082012.10.17
权利要求
1. 一种乙酰木聚糖酯酶,其特征在于其具有SEQ ID NO:2所示的氨基酸序列。
2. 根据权利要求1所述的乙酰木聚糖酯酶,其特征在于其蛋白质结构为同源六聚体或同源三聚体。
3. 包括权利要求1所述乙酰木聚糖酯酶的编码基因的表达载体。
4. 一种制备权利要求1所述的乙酰木聚糖酯酶的方法,其特征在于:用权利要求3所述的表达载体转化宿主菌,诱导表达获得重组乙酰木聚糖酯酶。
5. 权利要求1所述的乙酰木聚糖酯酶在水解短链脂肪酸脂、去酰基化反应、生产去酰基化的7-氨基头孢烷酸及去酰基化头孢菌素C中的应用。
说明书
技术领域
本发明属于基因工程技术领域,具体涉及一种从枯草芽孢杆菌(Bacillus subtilis)中获得的新型酯酶及表达载体、重组酯酶及重组酯酶在水解短链脂肪酸脂、去酰基化反应及β-内酰胺类抗生素的重要中间体-去酰基化的7-氨基头孢烷酸及去酰基化头孢菌素C中的应用。
背景技术
乙酰木聚糖酯酶(acetyl xylan esterase,缩写AXE)是一类可以将木聚糖或低聚木糖中的酰基化的吡喃木糖去酰基化并释放出醋酸的酯酶,分类于羧酸酯酶家族7,可以水解短链脂肪酸及各种酰基化化合物,具有广泛的底物特异性。
现今,乙酰木聚糖酯酶表现出巨大的科研价值和工业应用价值,主要体现在以下两个方面。一方面,乙酰木聚糖酯酶在半纤维素木聚糖的降解中起重要作用。该酶与内切β-1,4-木聚糖酶、β-D-木糖苷酶、α-L呋喃型阿拉伯糖苷酶及α-葡萄糖醛酸酶一起构成木聚糖降解酶系,能水解具有复杂结构的半纤维素木聚糖,使得开发和利用自然界中取之不尽的可再生资源成为可能,近些年获取高活力的乙酰基木聚糖酯酶及解析此酶的结构和功能已经成为研究的热点。另一方面,乙酰木聚糖酯酶具有良好的酰基酯酶活性,可以催化7-氨基头孢烷酸和头孢菌素C脱酰基合成β-内酰胺类抗生素母核,具有巨大的工业应用价值。β-内酰胺抗生素是抗生素中的最主要类型之一,全世界耗用量过万吨。目前使用的化学法存在成本高、三废污染严重等问题,很多制造领域已愈来愈趋向使用酶法合成。
目前已知报道的生产乙酰木聚糖酯酶的芽孢类微生物有Bacillus pumilus、Bacillus subtilis ATCC6633、Bacillus subtilis168、Bacillus halodurans C-125等。其中,除Bacillus subtilis ATCC6633的乙酰木聚糖酯酶被报道具有较高pNPA酶活力(1580 U/mg)外,其他菌株的乙酰木糖酯酶的活力和性能仍然较差,限制了其在半纤维素降解和酶法合成β-内酰胺抗生素中广泛应用。因此,需要开发一种高活性的乙酰木聚糖酯酶,扩大其使用范围。
发明内容
本发明的技术目的是提供一种新型的乙酰木聚糖酯酶的编码氨基酸序列及其对应的核苷酸序列。
本发明的第二个技术目的是提供乙酰木聚糖酯酶的蛋白质结构,其中,以同源六聚体的活性最高。
本发明的第三个技术目的是提供包括乙酰木聚糖酯酶的编码基因的表达载体。
本发明的第四个技术目的是提供制备重组乙酰木聚糖酯酶的方法。
本发明的第五个技术目的是提供重组乙酰木聚糖酯酶在水解短链脂肪酸脂、去酰基化反应、生产去酰基化的7-氨基头孢烷酸及去酰基化头孢菌素C中的应用。
本发明通过以下技术方案来实现:
本发明利用PCR技术从枯草芽孢杆菌基因组DNA中扩增出Cah基因,将Cah基因连在表达载体pET22b多克隆位点处,得到重组质粒pET-Cah,再将重组质粒pET-Cah转化E. coli BL21 (DE3),得到重组的大肠杆菌,即产乙酰木聚糖酯酶的基因工程菌E. coli BL21-pET-Cah。
菌株培养后用IPTG诱导表达,产生重组乙酰木聚糖酯酶,对重组乙酰木聚糖酯酶分离即可。详细的操作过程见实施例1和实施例2。
按照本发明的技术方案,可以达到良好的技术效果:
一、本发明的乙酰木聚糖酯酶序列与解淀粉芽孢杆菌FZB42(GenBank: YP 001419972.1)的乙酰木聚糖酯酶的序列同源性为99%,有10个碱基、2个氨基酸不同于解淀粉芽孢杆菌FZB42的乙酰基木聚糖酯酶基因:氨基酸位置第240位由GCC编码的丙氨酸(A)替换为ACC、或者ACA 、或者ACG编码的苏氨酸(T);289位氨基酸由GAA编码的谷氨酸(E)替换为CAA、或者CAG编码的谷氨酰胺(Q)。此外,本发明的乙酰木聚糖酯酶与已表征的Bacillus subtilis168(GenBank: NP_388200.1 )、Bacillus subtilis ATCC6633(GenBank: ZP_06875135.1 )、Bacillus pumilus Cect5072(GenBank: 2XLC_A)的乙酰木聚糖酯酶同源性分别为84%、84%、73%;与Thermobacillus composti乙酰木聚糖酯酶(GenBank: ZP_08919340.1)同源性为59%,与Thermotoga maritima MSB8乙酰木糖酯酶(GenBank: NP_227893.1)同源性41%,与 Bacillus halodurans C-125的头孢菌素C脱氢酶(GenBank: BAB07045.1)同源性35%。
对上述重组乙酰木聚糖酯酶进行了诱导表达,得到了如SEQ ID No:2所示的氨基酸序列。在本发明中,该氨基酸序列对应的核苷酸序列如SEQ ID No:1所示。但是,根据本领域技术人员的公知常识可知,可以编码一个氨基酸残基的核苷酸密码子的编码可能有至少一种,或者两种及以上,因此,本发明所述乙酰木聚糖酯酶的保护范围应该首先基于SEQ ID No:2所示的氨基酸序列为准,其扩展保护范围是可以编码出SEQ ID No:2所示的氨基酸序列的任何核苷酸序列,而不限于SEQ ID No:1所示的核苷酸序列所示的序列。
二、本发明所述的乙酰木聚糖酯酶(AXE),其蛋白质结构为同源六聚体,可解聚为同源三聚体,且同源三聚体具有一定的活性。
本发明的重组乙酰木聚糖酯酶单体分子量为36 kDa,而三聚体分子量为107 kDa,六聚体分子量为216 kDa。自然状态下乙酰木聚糖酯酶为同源六聚体,在1%的SDS作用下,可以解聚为同源三聚体,无SDS存在时,三聚体迅速自聚合为六聚体,且恢复六聚体的高活性。现有技术报道的Bacillus subtilis168及短小芽孢杆菌乙酰木聚糖酯酶自然状态下亦为同源六聚体,分子量分别为220 kDa和190 kDa,但其三聚体存在形式未见报道。(Multifunctional Xylooligosaccharide/Cephalosporin C Deacetylase Revealed by the Hexameric Structure of the Bacillus subtilis Enzyme at 1.9 A Resolution. Florence Vincent, Simon J et al. 2003. J. Mol. Biol. (2003) 330, 593-606;The crystal structure of the cephalosporin deacetylating enzyme acetyl xylan esterase bound to paraoxon explains the low sensitivity of this serine hydrolase to organophosphate inactivation. Silvia Montoro-Garcia, Fernando Gil-Ortiz et al. 2011.Biochem. J 436,321-330)。Bacillus subtilis ATCC6633的乙酰木聚糖酯酶为单聚体,分子量为37 kDa(YesT: A new rhamnogalacturonan acetyl esterase from Bacillus subtilis. Irene Martinez-Martinez, Jose Navarro-Fernandez et al. Proteins 2008; 71:379–388. )。Bacillus halodurans C-125的头孢菌素C脱氢酶为单聚体,分子量为45kDa(Characterization of a New C-125 Rhamnogalacturonan Acetyl Esterase from Bacillus halodurans with a New Putative Carbohydrate Binding Domain. Jose Navarro-Fernandez, Irene Martinez-Martinez et al. J. Bacteriol.2008, 190(4):1375.)。
本发明的来源于枯草芽孢杆菌的DNA序列在大肠杆菌体系中高效可溶的表达重组乙酰木聚糖酯酶具有广泛的底物谱,可以水解短链脂肪酸脂及多种酰基化化合物。本发明的重组乙酰木聚糖酯酶破胞后原始pNPA酶活力为383.3 U/mL;部分纯化后pNPA比活力达到2950U/mg,头孢菌素C的活力达到640 U/mg,均为芽孢杆菌属已知报道的最高活性,见表1。
本发明的重组乙酰木聚糖酯酶对C2的脂肪酸脂有非常强的链长专一性,链长增加为C4时活性已降低至0.09%,鲜见报道。与已知报道的Thermobifida fusca的乙酰木聚糖酯酶的比较见表2。
表2. 重组乙酰木聚糖酯酶对脂肪酸脂的链长专一性
[a], Yang, C.-H., & Liu, W.-H. (2008). Purification and properties of an acetylxylan esterase from Thermobifida fusca. Enzyme and Microbial Technology, 42(2), 181-186.
三、本发明还要求保护包括本发明所述乙酰木聚糖酯酶的编码基因的表达载体。根据本领域技术人员的公知常识,所述的表达载体应当包括但不限于重组载体、重组菌、转基因细胞系、表达盒或重组病毒;在本发明的实施例中,所述的具体的表达载体是为将本发明的乙酰木聚糖酯酶的编码基因插入载体pET22b的多克隆位点得到的重组载体。
四、一种制备本发明所述的乙酰木聚糖酯酶的方法,其特征在于:用本发明所述的表达载体转化宿主菌,诱导表达获得重组乙酰木聚糖酯酶。
五、本发明所述的乙酰木聚糖酯酶在水解短链脂肪酸脂、去酰基化反应、生产去酰基化的7-氨基头孢烷酸及去酰基化头孢菌素C中的应用。
本发明的应用表现为对目前7-氨基头孢烷酸和尤其是头孢菌素C的高酶活。Montoro-García已尝试对B. pumilus的乙酰木糖酯酶进行固定化,来催化合成7-ADCA和头孢菌素C,催化反应100min后转化率达到100%(A colorimetric assay for the determination of acetyl xylan esterase or cephalosporin C acetyl esterase activities using 7-amino phalosporanic acid, cephalosporin C, or acetylated xylan as substrate. Silvia Montoro-García, Fernando Gil-Ortiz et al. 2010. Bioresource Technology 101 331-336)。而本发明的重组乙酰木聚糖酯酶对7-氨基头孢烷酸和头孢菌素C的活性远远高于B. pumilus乙酰木聚糖酯酶,这显示出本发明乙酰木聚糖酯酶在β-内酰胺抗生素中间体的工业合成中具有巨大的应用潜力。
附图说明
图1为扩增出的Cah基因片段。其中,M: DNA Marker DL5000;1,2: Cah。
图2 为Cah和pET22b连接结果。其中,M: DNA Marker DL5000;1: pDK7-Nox。
图3为基因工程菌表达的乙酰木聚糖酯酶的SDS-PAGE凝胶(考马斯亮蓝染色);其中,M:宽分子量maker (TaKaRa),1-3 纯化的乙酰基木聚糖酯酶。
图4为牛血清蛋白(BSA)浓度-OD595吸光值标准曲线。
图5为对硝基苯酚(p-NP)浓度-OD405吸光值标准曲线。
图6 为醋酸浓度-OD210吸光值标准曲线。
图7 为纯化的乙酰木聚糖酯酶最适反应温度、pH及温度pH稳定性实验结果。
图8为非解聚与解聚的乙酰木聚糖酯酶的SDS-PAGE凝胶电泳(考马斯亮蓝染色);其中,M:宽分子量maker (TaKaRa),1、2分别为纯化的非解聚与解聚的乙酰基木聚糖酯酶。
图9为4-甲基伞形酮浓度-OD354的标准曲线。
图10为凝胶过滤分子量标准曲线。其中,Ve为样品洗脱体积,Vo为外水体积。
图11为SDS对乙酰木聚糖酯酶的影响。其中,泳道1为添加0.5%的SDS酶液,泳道2为不添加SDS的酶液;
图12为确定三聚体的活性及其与六聚体的变换关系。
本发明的乙酰木聚糖酯酶基因工程菌株,其分类命名为大肠杆菌(Escherichia coli)BL21-pET-Cah;其保藏机构全称为中国典型培养物保藏中心,简称CCTCC,地址为中国. 武汉. 武汉大学;保藏日期为2012年10月17日,保藏号编号为:CCTCC NO:M 2012408。
具体实施方式
本发明所使用的菌种来源:
1、表达菌E. coli BL21 (DE3)购买自TaKaRa公司。
2、枯草芽孢杆菌(Bacillus subtilis) CICC 20034的乙酰木聚糖酯酶的基因Cah来源:以Genebank公布的解淀粉芽孢杆菌FZB42中Cah基因(GenBank: ZP_10041520.1)为模板,自行设计引物,PCR扩增后得到。
3、所涉及引物为自行设计,并委托南京金斯瑞公司合成。
下面结合附图对本发明的技术方案做进一步的说明,但不应理解为对本发明的限制:
实施例1
本实施例说明产乙酰木聚糖酯酶的基因工程菌大肠杆菌(Escherichia coli)BL21-pET-Cah的构建。
本实施例所述的产乙酰木聚糖酯酶的基因工程菌大肠杆菌(Escherichia coli)BL21-pET-Cah,它是将枯草芽孢杆菌(Bacillus subtilis) CICC 20034的乙酰木聚糖酯酶的基因Cah通过表达载体pET22b转化E. coli BL21 (DE3) 得到的基因工程菌。
上述基因工程菌的构建方法为:利用PCR技术从质粒载体枯草芽孢杆菌基因组DNA中扩增出Cah基因,将Cah基因连在表达载体pET22b多克隆位点处,得到重组质粒pET-Cah,再将重组质粒pET-Cah转化E. coli BL21 (DE3),得到重组的大肠杆菌,即产乙酰木聚糖酯酶的基因工程菌E. coli BL21-pET-Cah。
上述构建方法,具体包括以下步骤:
(1) 基因Cah的克隆:
根据Genebank公布的解淀粉芽孢杆菌FZB42中Cah基因(GenBank: ZP_10041520.1),序列设计合引物:
P1:5’ TATACATATG(NdeI)CAATTATACGACTTGC 3’(SEQ ID NO:3)。
P2:5’ GGAATTC(EcoRI)CTCAGCCTTTCAGATGCGCT 3’(SEQ ID NO:4)。
引物两端分别引入限制性酶切位点NdeI和EcoRI,依照基因组提取试剂盒(OMEGA公司)的说明提取枯草芽孢杆菌基因组DNA,以枯草芽孢杆菌基因组DNA为模板完成PCR反应;
PCR的8管反应体系是:Buffer 22.5μL,ddH2O 141.3μL,MgCl2 13.5μL,dNTP-mix 18μL,引物P1 4.5μL,引物P2 4.5μL,Taq酶 2.7μL;混合均匀后,分成8管,再向每管加入2μL模板DNA;PCR反应条件:94 ℃预变性 4 min;进行30个循环反应:94 ℃变性45 s,54.1 ℃退火30 s,72 ℃延伸60 s;最后72 ℃保温10 min,反应结束后4 ℃保存,反应终止后,0.8 %琼脂糖凝胶电泳检测,结果如图1所示;经PCR纯化试剂盒(TaKaRa公司)纯化后,与pMD-T Vector载体连接,进行序列测定,NdeI和EcoRI酶切重组pMD-T Vector载体,回收957 bp片段,NdeI和EcoRI酶切pET22b载体,回收大片段产物,将双酶切产物在T4DNA连接酶作用下连接获得重组质粒pET-Cah。
(2) 重组基因工程菌的获得:
将重组质粒pET-Cah转化E. coli BL21 (DE3),涂布含40 μg/mL的氯霉素平板,挑取阳性转化子,并进行菌落PCR鉴定,得到重组E. coli,命名为E. coli BL21pET-Cah,提取E. coli BL21pET-Cah的重组质粒,进行NdeI和EcoRI双酶切验证(如图2)。
实施例2
本实施例说明乙酰木聚糖酯酶的诱导表达方法。
利用实施例1的基因工程菌诱导表达乙酰木聚糖酯酶的具体方案如下:
(1) 出发菌株:大肠杆菌(Escherichia coli)BL21-pET-Cah;
(2) 种子培养:
培养基:LB培养基:酵母粉 5g/L,蛋白胨10 g/L,氯化钠10 g/L;
培养条件:250 mL三角瓶,装液量50 mL,培养温度37 ℃,摇床转速200 r/min,培养12 h;
(3) 发酵培养:
培养基组成:LB培养基:酵母粉 5g/L,蛋白胨10 g/L,氯化钠10 g/L;
培养条件:接种量1.5% (v/v),发酵温度37 ℃,当OD达到0.6~0.8时加异丙基-β-D-硫代吡喃半乳糖苷(IPTG)诱导,IPTG的终浓度1.0 mmol/L,摇床转速180~250 r/min,发酵2-10 h。
(4) 粗酶液的获取:
取出发酵液,置于50mL 离心管中,4℃,10000转离心10min,弃去上清,加入50mL pH7.4的Tris-HCl缓冲液,对菌体进行超声破碎,破碎条件如下:3s 工作,3s间歇,550w 15min。破碎后4℃,10000转离心30min。上清即为粗酶液。
(5) 酶活力的测定:
使用pNPA为底物,1个酶活单位(U)定义为:在一定反应条件下,每分钟释放1μL p-NP所需的酶量。
标准曲线制作:A溶液的配制:50 mmol/L的磷酸缓冲(pH7.0),其中含有0.6% Triton X-100和0.1%的阿拉伯树胶;B溶液的配制:准确称取0.0139 g pNP标准物质,用A溶液溶解,容量瓶定容至250 mL,配成标准物质浓度为1 μmol/mL的B溶液。采用A溶液对B溶液进行梯度稀释,配成不同浓度的pNP溶液。在酶标板中加入10 μL缓冲液,然后依次加入不同浓度的pNP标准溶液各240 μL,对照中加入A溶液。用酶标仪测定405 nm波长处吸光值。以pNP的浓度作为横坐标,以吸光值作为纵坐标,作标准曲线(图5)。
测定步骤如下:在反应体系中加入240 μL 底物溶液(A溶液与B溶液按9:1混合),10 μL 适当稀释的酶液,置于水浴锅中,温度为40 ℃,反应时间10 min;使用酶标仪在410 nm处测定吸光值。实验设三个平行,取平均值,并使用10 μL 蒸馏水最为对照组。p-NP标准曲线及方程如图5。
测定结果显示:100倍稀释的酶液,A410nm下吸光值分别为:1.449、1.482、1.509,通过标准曲线(图5)计算,对应活性为224.96U/mL。
(6) 蛋白量的测定
使用Brandford法测定蛋白量。
蛋白标曲的制作:配制考马斯亮蓝溶液:100 mg 考马斯亮蓝G-250溶于50 mL 95 % 乙醇中,再加入100 mL 85 %(v/v) H3PO4,最后用蒸馏水稀释至1 l,滤纸过滤后可使用;配制0.1 g/l BSA:称取0.01 g BSA,溶于10 mL 蒸馏水中,使用前用生理盐水配制成0.01、0.02 、0.03、0.04、0.06、0.08 mg/mL;取7个2 mL离心管,编好号(0,1,2,3,4,5,6),在1~6号离心管中分别加入上述稀释后各浓度BSA溶液0.3 mL,最后在每支离心管中加入1.2 mL 考马斯亮蓝溶液。在0号离心管中1.2 mL 考马斯亮蓝溶液和0.3 mL生理盐水。各管混合后后,以0号管为对照测定A595。测得的标准曲线如图4。
蛋白的检测具体步骤如下:首先以牛血清蛋白(BSA)标样的浓度对OD595吸光值作标准曲线方程,如图4。然后去适当稀释的酶液300μL ,加入1.2mL考马斯亮蓝工作液,混匀,室温放置15min,与595nm下测定吸光值。每组做两个平行样,以蒸馏水为空白。
测定结果显示稀释5倍的酶液反应后的OD595吸光值为,0.252、0.277。按照图4的标准曲线计算后,蛋白浓度为:0.16 mg/μL 。因此,工程菌发酵液的乙酰基木聚糖酯酶粗酶液的比酶活力为 1406 U/mg。
实施例3
本实施例说明乙酰木聚糖酯酶的纯化、分子量及其多聚体形式。
1. 乙酰木聚糖酯酶的纯化:
通过50%-80%(w/v)硫酸铵分级沉淀和超滤乙酰木聚糖酯酶进行浓缩和纯化。纯化步骤如下:
(1)50%-80% 硫酸铵分级沉淀:取破碎好的溶液,12000g,30min 离心;转移50mL上清至100mL烧杯中,边搅拌边慢慢加入50mL的SAS溶液(饱和硫酸铵溶液),将溶液放在磁力搅拌器上搅拌6小时后,放置于4度冰箱过夜充分沉淀。蛋白质溶液10000g 离心 30min。将50mL上清转移至新的烧杯,边搅拌边慢慢加入75mL的SAS溶液,使终浓度为80%的硫酸铵溶液,将溶液放在磁力搅拌器上搅拌6小时后,放置于4度冰箱过夜充分沉淀。蛋白质溶液10000g 离心 30min后,弃滤液,将沉淀用5mL pH7.0的磷酸缓冲溶液溶解,重新溶解后的溶液转移至透析袋中12h (4℃) 后,置于500mL的透析缓冲中,每3h换透析缓冲液一次,以彻底除去硫酸铵。测定纯化后的蛋白质含量及酶活力。
(2) 凝胶过滤:采用凝胶过滤对浓缩后的乙酰木聚糖酯酶进行进一步纯化,使用HiLoad 16/60 Superdex 200凝胶色谱柱(Healthcare Bio-Sciences 公司), 50 mM Tris-HCl (包含100 mM KCl,pH 7.5) 作为流动相,上样量100 μL,流速0.1 mL/min, 280nm检测。使用自动馏分收集器收集洗脱液,对有紫外吸收峰值的洗脱液进行酶活力测定,将有活力的部分合并,超滤浓缩后,即为纯酶液。测定纯化后的蛋白质含量及酶活力。
纯化效果如表3所示,纯化后的乙酰木聚糖酯酶显示出单一条带,如图3。经纯化后的乙酰基木糖酯酶经SDS电泳得到单一蛋白条带。这表明,SEQ ID NO.1所述核苷酸序列所编码的多肽在大肠杆菌E. coli BL21 (DE3) 中得到高效表达,且所有的重组蛋白均是可溶的,无包涵体形成。
2. 乙酰木聚糖酯酶分子量及其多聚体形式
采用凝胶过滤确定乙酰木聚糖酯酶自然状态下的分子量,操作流程如凝胶过滤的纯化流程。凝胶过滤蛋白标准(购自SIGMA, 货号:MWGF 1000),标曲的制作按照蛋白标准说明书,标曲见表图10。凝胶过滤结果显示, V0为43.49 mL,Ve为61.76 mL(Ve为紫外吸收峰出现时使用的流动相体积),未检测到其他紫外吸收峰。依照标曲计算得到乙酰木聚糖酯酶自然状态下的分子量为216 kDa。采用SDS-PAGE电泳(12%)乙酰木聚糖酯酶单亚基的分子量,SDS-PAGE凝胶的配制如表4,使用Bio-Rad 电泳仪,电泳条件为: 80V 45 min; 120V 45min。电泳样品经SDS-PAGE上样缓冲处理后煮沸,蛋白亚基彻底解聚。结果显示其分子量为36 kDa,如图8,泳道2。这表明乙酰木聚糖酯酶天然状态下为六聚体形式。
3. 乙酰木聚糖酯酶的三聚体
采用SDS-PAGE电泳(12%)对未解聚的乙酰木聚糖酯酶(酶液经SDS-PAGE上样缓冲处理后40 ℃温浴5 min,蛋白分子未解聚)进行分析,其他操作步骤如上述SDS-PAGE电泳。结果显示,乙酰木聚糖酯酶分子量为107 kDa(如图8,泳道1)。这表明乙酰木聚糖酯酶电泳后以三聚体形式存在,可能电泳过程中某些条件或电泳溶液中的某个物质使六聚体解聚为三聚体。
SDS是一种螯合剂,可以引起蛋白质变性,可能会引起六聚体的解聚。因此,为验证是否因电泳液中的SDS使天然六聚体解聚为三聚体,采用非变性聚丙烯酰胺凝胶电泳和酯酶染色进行检测。实施过程如下:
溶液的配制:凝胶的配制如表4(不添加SDS);Tris–甘氨酸电泳缓冲液:25 mmol/L Tris;250 mmol/L甘氨酸 (pH 8.3);样品缓冲液(0.1 mol/L Tris-Hcl,pH 6.8)2*的上样缓冲:0.5 mL 1 mol/L的Tris缓冲(PH 6.8)中加入1 mL甘油和0. 0001 g溴酚蓝,灭菌的双蒸水稀释至10 mL。处理样品后,加入SDS,使其终浓度为0.5 %,40℃ 温浴2 h后进行电泳,以不加SDS的酶液样品为空白。使用Bio-Rad 垂直电泳仪80 V电泳 45 min、120 V 电泳45 min;之后进行酯酶活性染色。活性染色的具体步骤如下:准确称取100 mg α-醋酸萘酯和100 mg β-醋酸萘酯,在2 mL丙酮中完全溶解,加入100 mg坚牢蓝,混合均匀。用0.1 mol/L pH 7.0磷酸缓冲液定容至100 mL;将凝胶放进染色液中,置于摇床上缓慢摇动,37 ℃,55转,染色2 min。结果如图11所示,未经SDS处理的空白样品为一条酯酶条带,而加入SDS的乙酰木聚糖酯酶在正常酯酶条带下方出现另外一条酯酶条带。这说明是SDS使乙酰木聚糖酯酶由天然的六聚体解聚成为三聚体,且0.5%的SDS不足以使六聚体完全解聚。加大SDS的浓度为1%,处理酶液2 h,进行聚丙烯酰胺凝胶电泳和酯酶染色,结果如图12,1%的SDS作用2 h可以使六聚体基本完全解聚为三聚体(如图12,泳道1)。
为确定三聚体的活性及其与六聚体的变换关系,在酯酶染色液中加入1%的SDS进行染色,以保证酯酶染色过程中,乙酰木聚糖酯酶始终以三聚体存在,从而确定三聚体的酯酶活性。结果显示,乙酰木聚糖酯酶的三聚体具有一定的活性,但低于六聚体(如图12,泳道2)。将凝胶转移至不含SDS的染色液中进行染色,结果显示乙酰木聚糖酯酶短时间内(1 min)恢复为高活性,如图12,泳道3。这表明,SDS可以使乙酰木聚糖酯酶解聚暂时为三聚体,洗去SDS后,乙酰木糖酯酶在短时间内迅速自聚合为六聚体。
表4. Native 聚丙烯酰胺电泳凝胶的配制
实施例4
本实施例说明重组乙酰木聚糖酯酶的基本酶学性质研究。
1. 乙酰木聚糖酯酶的最适反应温度、最适pH:
以4-甲基伞形乙酯为底物,检测乙酰木聚糖酯酶的最适反应温度、最适pH。1个酶活单位(U)定义为:在一定反应条件下,每分钟释放1 μL 4-甲基伞形酮所需的酶量。
具体步骤如下:
(1) 4-甲基伞形乙酯酶活力标曲的制作:配制0.1、0.15、0.2、0.25、0.3、0.35、0.4 mM的4-甲基伞形酮,分别测量其在354 nm下的吸光值。结果如图9。
(2) 最优反应温度的测定:混合100 μL pH 7.0的磷酸缓冲液(0.1 mol/L)与280 μL蒸馏水,40 ℃预热10 min;加入10 μL酶液;加入10 μL 100 mol/L 4-甲基伞形乙酯(溶解于DMSO);在30、40、45、50、55、60、70℃ 下反应10 min后,加入600 μL 50 mmol/L 柠檬酸,测354 nm下的吸光值。按照图9的标准曲线换算为酶活力。结果如图7(a)所示,最优反应温度为50℃。
(3) 最优反应pH的测定:混合100 μL pH分别为 3.0、4.0、5.0、6.0、7.0、7.5、8.0、8.5、9、10、11、12的磷酸缓冲液(0.1 mol/L)与280 μL蒸馏水,40℃预热10 min;其他步骤如(2)中所述。按照图9的标准曲线换算为酶活力。结果如图7(c)所示,最优反应pH为8.0。
2. 乙酰木聚糖酯酶的温度、pH稳定性:
使用pNPA为底物测定乙酰木聚糖酯酶的温度、pH稳定性。
(1)温度稳定性测定步骤:准备6个2 mL离心管,分别加入2 mL纯化的酶液;置于25-70℃水浴中,分别在0.5h、1 h、2 h、3 h、4 h、5 h、6 h、9 h、12 h、24 h测残余pNPA酶活力,并计算相对活力。结果如图7(b)所示,重组的乙酰木聚糖酯酶在55 ℃以下酶活力保持在80 %以上长达5天,高于60 ℃时仅可稳定4 h。
(2) pH稳定性测定步骤:准备10个2 mL离心管,放入1.9 mL pH 3-12的缓冲液,分别加入100 μL酶液。40 ℃水浴1 h后测残余酶活,并计算性对活力。结果如图7(b)所示,重组的乙酰木聚糖酯酶在pH 7.0-11.0范围内很稳定,酸性条件下不稳定。
实施例5
本实施例说明乙酰木聚糖酯酶对短链脂肪酸脂及酰基化合物的水解活性。
本研究考察了乙酰木聚糖酯酶在水解短链脂肪酸脂及去酰基化类反应(对硝基苯酚乙酸酯、对硝基苯酚丁酸酯、五酰基葡萄糖、4-甲基伞形乙酯、α- 醋酸萘酯、β- 醋酸萘酯)中的应用。具体实施方案如下:
1. 对硝基苯酚乙酸酯、对硝基苯酚丁酸酯的酶活测定:如实施案例2中酶活力的测定。结果见表5。
2. 对4-甲基伞形乙酯的酶活力测定:如实施案例4中的实验步骤。结果见表5。
3. 对五酰基葡萄糖的酶活力测定:1个酶活单位(U)定义为:在一定反应条件下,每分钟释放1 μL醋酸所需的酶量。醋酸的浓度由高效液相色谱法(日本到岛津公司,C18柱)进行检测。检测条件如下:流速0.6 mL/min,93.5 %的0.1 %磷酸,3.5 %的乙腈,紫外方法检测。
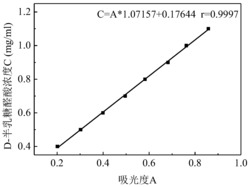
(1) 醋酸浓度标准曲线的制作:配制0.1、0.2、0.4、0.6 g/L的醋酸溶液,经液相检测210 nm下的吸光值。结果如图6所示。
(2) 对五酰基葡萄糖的酶活力测定:50 ℃ 预热490 μL 2 mmol/L 的五酰基葡萄糖溶液(溶于pH 8.0 的Tris-HCl缓冲液) 10 min;加入10 μL 酶液,空白组为蒸馏水,混匀后50 ℃水浴10 min;加入100 μL 0.33 mol/L 硫酸溶液,终止反应;高效液相色谱法分析醋酸含量。按图6所示标准曲线测定乙酰木聚糖酯酶对五酰基葡萄糖的活力。
结果为:两个平行的50倍稀释的乙酰木聚糖酯酶反应后溶液在210 nm检测下醋酸的峰面积为0.504、0.468 mAU*min,取平均值 0.486 mAU*min,按照6所示标准曲线计算乙酰木聚糖酯酶对五酰基葡萄糖的活力为328.5 U/mL,比活力为896U/mg。
4. 对α- 醋酸萘酯、β- 醋酸萘酯的酶活力测定
对α- 醋酸萘酯、β- 醋酸萘酯的酶活力测定的具体步骤如下:
(1) 各溶液的配制:A 底物液:2 mmol/ L α- 醋酸萘酯,称取20 mg α- 醋酸萘酯 (化学纯) 用1 mL 异丙醇溶解后加0. 1 mol/ L pH 7.4磷酸盐缓冲液 49 mL,用力振摇助溶,4 ℃ 避光保存可用1周;0. 1 mol/ L pH 7.4磷酸盐缓冲液:1.36 g KH2PO4溶于80 mL双蒸水中,用1 mol/L NaOH调pH 至7.4,定容至100 mL;B 终止液:1. 75 mol/L 醋酸(11 mL冰醋酸稀释至100mL);C 显色液:1 mmol/ L 固蓝B 盐,称取50 mg 固蓝B (分析纯) 加 0. 8% 吐温-20 100 mL,4 ℃避光保存可用 2周;D标准液:2 mmol/ L α- 萘酚,称取28. 8 mg α- 萘酚 ( 分析纯) 加无水乙醇 0. 5 mL 溶解后水补足至100 mL,4 ℃ 避光保存。
(2) 操作步骤:取3个1.5 mL离心管,分别标明测定、标准、空白。分别向离心管中加入0.5 mL底物液,50 μL待测样品或标准液或蒸馏水,40 ℃ 水浴20 min;分别取出250 μL反应后的溶液,加入0.5 mL终止液和0.5 mL显色液,混匀,40 ℃ 水浴20 min;
以空白管调零,520 nm下读取吸光度。
(3) 酶活力的计算:测定管A /标准管A*100 = α/β-醋酸萘酯酶酶活力(U/L);酶活力定义:每升酶液在40 ℃ 酶反应 1 min 产生1 mol α/β- 萘酚为 1 个酶活性单位(U/ L)。
测得的α/β-醋酸萘酯的活力见表5。
表5. 乙酰木聚糖酯酶对短链脂肪酸脂及酰基化合物的水解活性
实施例6
本实施例说明重组乙酰木聚糖酯酶在生产β-内酰胺类抗生素的重要中间体---去酰基化的7-氨基头孢烷酸中的应用。
乙酰木聚糖酯酶可利用7-氨基头孢烷酸为原料,经过去酰基化转化为β-内酰胺类抗生素的重要中间体---7-氨基头孢烷酸。本发明初步探究了此新颖的乙酰木聚糖酯酶对7-氨基头孢烷酸的去酰基化酶活力。1个酶活单位(U)定义为:在一定反应条件下,每分钟释放1 μL醋酸所需的酶量。
具体操作步骤如下:
(1) 醋酸浓度标准曲线的制作:如实施案例5中醋酸浓度标准曲线的制作。
(2) 7-氨基头孢烷酸酶活力的测定:50 ℃ 预热490 μL 2 mmol/L 的7-氨基头孢烷酸溶液(溶于pH 8.0 的Tris-HCl缓冲液) 10 min;加入10 μL 酶液,空白组为蒸馏水,混匀后50 ℃水浴10 min;加入100 μL 0.33 mol/L 硫酸溶液,终止反应;高效液相色谱法分析醋酸含量。按图6所示标准曲线测定乙酰木聚糖酯酶对五酰基葡萄糖酶的活力。
结果为:两个平行的10倍稀释的乙酰木聚糖酯酶反应后溶液在210 nm检测下醋酸的峰面积为1.556、1.506 mAU*min,取平均值 1.531 mAU*min,按照6所示标准曲线计算乙酰木聚糖酯酶对五酰基葡萄糖酶的活力为135.6 U/mL,比活力为638.7U/mg。
实施例7
本实施例说明重组酯酶在生产β-内酰胺类抗生素的重要中间体---去酰基化头孢菌素C中的应用。
乙酰木聚糖酯酶可利用头孢菌素C为原料,经过去酰基化转化为β-内酰胺类抗生素的重要中间体—去酰基化的头孢菌素C。本发明探究了此酶对头孢菌素C的去酰基化酶活力。1个酶活单位(U)定义为:在一定反应条件下,每分钟释放1 μL醋酸所需的酶量。
具体步骤同实施案例7,结果为:两个平行的10倍稀释的乙酰木聚糖酯酶反应后溶液在210 nm检测下醋酸的峰面积为1.417、1.406 mAU*min,取平均值 1.4115 mAU*min,按照6所示标准曲线计算乙酰木聚糖酯酶对7-氨基头孢烷酸的活力为127.6 U/mL,比活力为348 U/mg。
综上所述,本发明的来源于枯草芽孢杆菌的DNA序列在大肠杆菌体系中高效可溶的表达重组乙酰木聚糖酯酶具有广泛的底物谱,可以水解短链脂肪酸脂及多种酰基化化合物,且高活性催化合成7-氨基头孢烷酸及去酰基化的头孢菌素C,在木聚糖降解及β-内酰胺类抗生素生产的应用方面具有巨大的应用潜力。
上述实施例为本发明较佳的实施方式,但本发明的实施方式并不受上述实施例限制,其他的任何未背离本发明的精神实质与原理下所作的改变、修饰、替代、组合、简化均应为等效的置换方式,都包含在本发明的保护范围。
序列表
<110> 南京工业大学
<120> 一种乙酰木聚糖酯酶及其应用
<130> NJUT121018
<160> 4
<170> PatentIn version 3.3
<210> 1
<211> 957
<212> DNA
<213> Bacillus subtilis
<220>
<221> CDS
<222> (1)..(957)
<400> 1
atg caa tta tac gac ttg ccg ctc gac caa ctg caa acc tat aaa cca 48
Met Gln Leu Tyr Asp Leu Pro Leu Asp Gln Leu Gln Thr Tyr Lys Pro
1 5 10 15
aat aaa acc gct cct cat gac ttt tcg gat ttt tgg gca tca tca tta 96
Asn Lys Thr Ala Pro His Asp Phe Ser Asp Phe Trp Ala Ser Ser Leu
20 25 30
cat gag ctt gca aag gaa gag gca aag cct gaa ttg aag gct gaa agc 144
His Glu Leu Ala Lys Glu Glu Ala Lys Pro Glu Leu Lys Ala Glu Ser
35 40 45
tac ccg gct gac ggc gtg aaa gta ttc cgg ctg aca tac cgt agc ttc 192
Tyr Pro Ala Asp Gly Val Lys Val Phe Arg Leu Thr Tyr Arg Ser Phe
50 55 60
ggc aag gcg gaa att gag gga tgg tac gcc gtt ccc gac cgg caa gga 240
Gly Lys Ala Glu Ile Glu Gly Trp Tyr Ala Val Pro Asp Arg Gln Gly
65 70 75 80
ccg cat ccg gcc atc gtg aaa tat cac ggc tat aac gcg agc tat gac 288
Pro His Pro Ala Ile Val Lys Tyr His Gly Tyr Asn Ala Ser Tyr Asp
85 90 95
ggg gat ata cat gac atc gta aat tgg gcg ctt cac gga tat gcc gcg 336
Gly Asp Ile His Asp Ile Val Asn Trp Ala Leu His Gly Tyr Ala Ala
100 105 110
ttc ggc atg ctt gtc cgc ggc cag cac agc agc aca gat aca agc gtc 384
Phe Gly Met Leu Val Arg Gly Gln His Ser Ser Thr Asp Thr Ser Val
115 120 125
tca ccg cac gga cac gtt ccc ggc tgg atg acg aaa ggg att tta gat 432
Ser Pro His Gly His Val Pro Gly Trp Met Thr Lys Gly Ile Leu Asp
130 135 140
aaa gat aca tat tat tac cgc ggc gtg tac tta gac gcc gtc cgg gct 480
Lys Asp Thr Tyr Tyr Tyr Arg Gly Val Tyr Leu Asp Ala Val Arg Ala
145 150 155 160
ttg gaa gtc atc agc ggc ttt gat gaa gtg gat gaa acg agg atc gcc 528
Leu Glu Val Ile Ser Gly Phe Asp Glu Val Asp Glu Thr Arg Ile Ala
165 170 175
gtc atc ggc ggg agc cag gga ggc gga ctt tca atc gct gcg gcg gca 576
Val Ile Gly Gly Ser Gln Gly Gly Gly Leu Ser Ile Ala Ala Ala Ala
180 185 190
cta tct gat att ccg aga gcg gtt gcg gcc gat tat cct tat tta agc 624
Leu Ser Asp Ile Pro Arg Ala Val Ala Ala Asp Tyr Pro Tyr Leu Ser
195 200 205
aat ttt gag cgg gcc att gat gtt gct ctt gat gag ccg tat ctt gag 672
Asn Phe Glu Arg Ala Ile Asp Val Ala Leu Asp Glu Pro Tyr Leu Glu
210 215 220
att aac tcc ttt ttc aga aag aac ggc agc cct gaa acg gag aag acc 720
Ile Asn Ser Phe Phe Arg Lys Asn Gly Ser Pro Glu Thr Glu Lys Thr
225 230 235 240
gcg atg aat aca ctg gcc tat ttt gat atc atg aat ctg gcg gac cgc 768
Ala Met Asn Thr Leu Ala Tyr Phe Asp Ile Met Asn Leu Ala Asp Arg
245 250 255
gta aaa gtt ccg gtg ctc atg tcc atc ggg ctg att gac cgc gtg act 816
Val Lys Val Pro Val Leu Met Ser Ile Gly Leu Ile Asp Arg Val Thr
260 265 270
ccg ccg tcg acg gtg ttt gcc gct tac aat cat ctg gaa acg gaa aaa 864
Pro Pro Ser Thr Val Phe Ala Ala Tyr Asn His Leu Glu Thr Glu Lys
275 280 285
caa ttg aag gtt tac cgg tat ttc gga cat gaa tac ata cct tct ttc 912
Gln Leu Lys Val Tyr Arg Tyr Phe Gly His Glu Tyr Ile Pro Ser Phe
290 295 300
cat acg gaa aag ctg gcc ttt ctg aaa gcg cat ctg aaa ggc tga 957
His Thr Glu Lys Leu Ala Phe Leu Lys Ala His Leu Lys Gly
305 310 315
<210> 2
<211> 318
<212> PRT
<213> Bacillus subtilis
<400> 2
Met Gln Leu Tyr Asp Leu Pro Leu Asp Gln Leu Gln Thr Tyr Lys Pro
1 5 10 15
Asn Lys Thr Ala Pro His Asp Phe Ser Asp Phe Trp Ala Ser Ser Leu
20 25 30
His Glu Leu Ala Lys Glu Glu Ala Lys Pro Glu Leu Lys Ala Glu Ser
35 40 45
Tyr Pro Ala Asp Gly Val Lys Val Phe Arg Leu Thr Tyr Arg Ser Phe
50 55 60
Gly Lys Ala Glu Ile Glu Gly Trp Tyr Ala Val Pro Asp Arg Gln Gly
65 70 75 80
Pro His Pro Ala Ile Val Lys Tyr His Gly Tyr Asn Ala Ser Tyr Asp
85 90 95
Gly Asp Ile His Asp Ile Val Asn Trp Ala Leu His Gly Tyr Ala Ala
100 105 110
一种乙酰木聚糖酯酶及其应用专利购买费用说明
![]()
Q:办理专利转让的流程及所需资料
A:专利权人变更需要办理著录项目变更手续,有代理机构的,变更手续应当由代理机构办理。
1:专利变更应当使用专利局统一制作的“著录项目变更申报书”提出。
2:按规定缴纳著录项目变更手续费。
3:同时提交相关证明文件原件。
4:专利权转移的,变更后的专利权人委托新专利代理机构的,应当提交变更后的全体专利申请人签字或者盖章的委托书。
Q:专利著录项目变更费用如何缴交
A:(1)直接到国家知识产权局受理大厅收费窗口缴纳,(2)通过代办处缴纳,(3)通过邮局或者银行汇款,更多缴纳方式
Q:专利转让变更,多久能出结果
A:著录项目变更请求书递交后,一般1-2个月左右就会收到通知,国家知识产权局会下达《转让手续合格通知书》。













动态评分
0.0