

IPC分类号 : C12N9/80,C12N15/55,C12N15/63,C12N5/10,C12N1/21,C12P35/00,C12R1/19





专利摘要
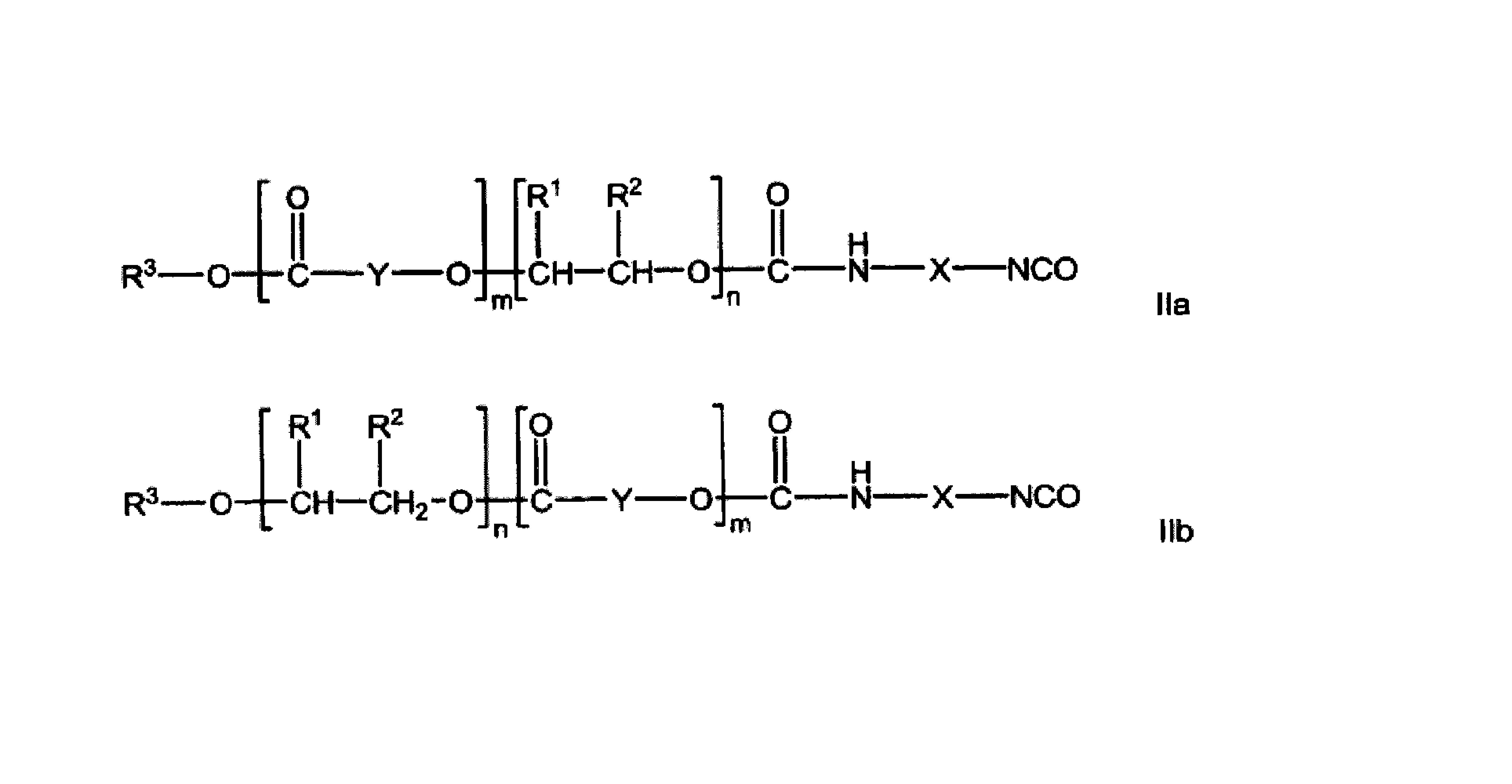
本发明公开了一种头孢菌素酰化酶突变体及其编码基因与应用。本发明提供了的蛋白,为如下1)-3)中的任一一种:1)序列表中序列4所示的氨基酸序列组成的蛋白;2)序列表中序列3所示的氨基酸序列组成的蛋白;3)将序列表中序列3或序列4的氨基酸残基序列经过一个或几个氨基酸残基的取代和/或缺失和/或添加且具有头孢菌素酰化酶功能的由1)衍生的蛋白。本发明的实验证明,本发明提供了一种野生型CPC酰化酶变体,通过HPLC结果表明本发明的野生型CPC酰化酶变体与野生型酶相比,对CPC的比活可提高6.5倍。在一步法转化中,在3h内,转化率达到98%以上。
说明书
技术领域
本发明涉及生物技术领域,尤其涉及一种头孢菌素酰化酶突变体及其编码基因与应用。
背景技术
头孢菌素类药物(Cephalosporins)是一类重要的β-内酰胺类半合成广谱抗生素。目前工业上采用发酵技术生产头孢菌素C(Cephalosporin C,缩写为CPC),经过化学法裂解或酶法裂解,产生重要的中间体化合物7-氨基头孢烷酸(7-aminocephalospora-nic acid,缩写为7-ACA)。再以7-氨基头孢烷酸作为母核,与不同的侧链缩合,得到半合成的β-内酰胺类抗生素。目前化学法生产7-氨基头孢烷酸因为造成环境污染,已经逐渐被淘汰,取而代之的是利用酶法进行生产。酶法生产主要分为两步酶法和一步酶法。两步酶法首先利用D-氨基酸氧化酶(D-amino acid oxidase),生成中间物戊二酰-7-氨基头孢烷酸(Glutaryl-7-aminocephalosporanic acid,缩写为Gl-7ACA),之后通过头孢菌素酰化酶生成最终产物7-氨基头孢烷酸。而一步酶法仅需头孢菌素酰化酶就可以将头孢菌素C直接转化为7-氨基头孢烷酸,优于两步酶法。
头孢菌素酰化酶的主要底物为GL-7ACA,而对CPC活性较低。根据同源性,可以将其分为5类。2000年,Kim等人(Kim,Y.,et al.,The 2.0 crystal structure of cephalosporin acylase.Structure,2000.8(10):p.1059-1068.)解出第一个头孢菌素酰化酶的晶体结构CAD。来源于假单胞菌(Pseudomonas diminuta KAC-1),属于第一类的头孢菌素酰化酶,此后CAD与底物GL-7ACA共结晶的结构和来源于Pseudomonas sp.GK16,同属第一类的结构也相继解出。晶体结构的出现为进一步认识和改造提供了条件。在五类酰化酶中,第三类催化头孢菌素C的活性最高,为催化GL-7ACA活性的4%(Aramori,I.,et al.,Comparative characterization of new glutaryl 7-ACA and cephalosporin C acylases.Journal of Fermentation and Bioengineering,1992.73(3):p.185-192)。
发明内容
本发明的一个目的是提供一种头孢菌素酰化酶突变体或其功能等效衍生物。
本发明提供的头孢菌素酰化酶突变体或其功能等效衍生物,其特征在于:将野生型头孢菌素酰化酶的氨基酸序列进行如下至少一种氨基酸取代得到的蛋白:1)自N末端第296位(β亚基的57位)的组氨酸(H)被丙氨酸(A)所取代;2)自N末端第309位(β亚基的70位)的组氨酸(H)被缬氨酸(Y)所取代;
所述野生型头孢菌素酰化酶的氨基酸序列为序列表中的序列1。
本发明的另一个目的是提供的蛋白,为如下1)-3)中的任一一种:
1)序列表中序列4所示的氨基酸序列组成的蛋白;
2)序列表中序列3所示的氨基酸序列组成的蛋白;
3)将序列表中序列3或序列4的氨基酸残基序列经过一个或几个氨基酸残基的取代和/或缺失和/或添加且具有头孢菌素酰化酶功能的由1)衍生的蛋白。
所述一个或几个氨基酸残基的取代和/或缺失和/或添加为不超过10个氨基酸残基的取代和/或缺失和/或添加。具体为3)所示蛋白的氨基酸序列为序列表中的序列8。
所述头孢菌素酰化酶突变体或其功能等效衍生物或所述蛋白的编码基因也是本发明保护的范围。
所述的编码基因为如下1)-5)中任一所述的DNA分子:
1)序列表中的序列6所示的DNA分子;
2)序列表中的序列5所示的DNA分子;
3)序列表中的序列7所示的DNA分子;
4)在严格条件下可与1)或2)或3)限定的DNA序列杂交且编码具有头孢菌素酰化酶功能的蛋白的DNA分子;
5)与1)或2)或3)限定的DNA序列具有90%以上的同源性且编码具有头孢菌素酰化酶功能的蛋白的DNA分子。
上述严格条件可为在0.1×SSPE(或0.1×SSC),0.1% SDS的溶液中,在65℃条件下杂交并洗膜。
含有所述编码基因的重组载体、表达盒、转基因细胞系或重组菌也是本发明保护的范围。
所述重组载体为将所述头孢菌素酰化酶突变体或其功能等效衍生物或所述蛋白的编码基因插入pET30(a)的Xba I和Xho I酶切位点间,得到表达所述头孢菌素酰化酶突变体或其功能等效衍生物或所述的蛋白的载体,也可插入pET30(a)的Nde I和Xho I酶切位点间,得到同样的载体。
所述重组菌为将所述的重组载体转入宿主菌中,得到的重组菌,所述重组菌具体为大肠杆菌。
所述头孢菌素酰化酶突变体或其功能等效衍生物或所述的蛋白、所述编码基因或所述的重组载体、表达盒、转基因细胞系或重组菌在作为头孢菌素酰化酶中的应用也是本发明保护的范围。
本发明的第三个目的是提供一种制备头孢菌素酰化酶的方法。
本发明提供的方法,包括如下步骤:发酵所述的重组菌,收集发酵产物,即得到头孢菌素酰化酶。
所述头孢菌素酰化酶突变体或其功能等效衍生物或所述的蛋白、所述编码基因或所述的重组载体、表达盒、转基因细胞系或重组菌在制备7-氨基头孢烷酸中的应用也是本发明保护的范围。
本发明的实验证明,本发明提供了野生型CPC酰化酶变体,通过HPLC结果表明本发明的野生型CPC酰化酶变体与野生型酶相比,对CPC的比活可提高6.5倍。在一步法转化中,在3h内,转化率达到98%。
附图说明
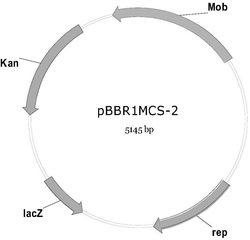
图1为重组质粒pET30a-CA的物理图谱
图2为突变株CA野生型,CA-113,CA-1C,阴性对照的SDS-PAGE电泳图片
图3为突变株CA-1C催化底物CPC生成产物7-ACA的HPLC峰图
A为反应初始时体系的HPLC峰图
B为反应结束时体系的HPLC峰图
C为7-ACA的标准品HPLC峰图
图4为突变株CA-1C催化底物CPC转化率曲线
具体实施方式
下述实施例中所使用的实验方法如无特殊说明,均为常规方法。
下述实施例中所用的材料、试剂等,如无特殊说明,均可从商业途径得到。
实施例1、头孢菌素酰化酶突变体的获得
一、含有野生型头孢菌素酰化酶CA的表达载体pET30(a)-CA-WT
通过NCBI(www.ncbi.nlm.nih.gov/)查询得到Pseudomonas SE83 acyII的头孢菌素酰化酶的氨基酸序列(GenBank:AAA25690.1,氨基酸序列为序列1),采用重叠PCR的方法,合成适合于在E.coli中表达的头孢菌素酰化酶基因的核苷酸序列。具体是将其全长的氨基酸序列在线输入DNAworks程序(http://helixweb.nih.gov/dnaworks/)。通过设定密码子偏爱性,DNAWorks输出适合于在E.coli中表达的头孢菌素酰化酶基因的核苷酸序列片段,每条长度约为45bp共78条相互重叠的寡核苷酸序列。将合成的寡聚核苷酸片段用10mM Tris-HCl,pH 8.0的缓冲液溶解。PCR反应体系为在0.5mlPCR薄壁管,加入无菌水6.3μl、按照每条寡核苷酸片段终浓度25nm、10μl 2×GC bufferI、3.2μl四种核苷酸混合液(浓度为2.5mM),0.5μl Takara公司的La-taq聚合酶,体系共20μl。反应条件为,先94℃ 2分钟;然后94℃ 1分钟,63℃ 1分钟,72℃ 5分钟,共29个循环;最后72℃ 10分钟。最后使用含有Nde I酶切位点的上游引物CA-1:
5’-GGGACACCATATGACCATGGCGGCGAAAACC-3’(带下划线碱基为限制性内切酶Nde I识别位点)和含有BamH I识别位点的下游引物CA-2:
5’-CTCGCGGGATCCTTACGCCGGCACCAGTTCCTG-3’(带下划线碱基为限制性内切酶BamH I识别位点)。PCR反应体系为在0.5mlPCR薄壁管,依次加入2.7μl的无菌水、10μl 2×GC bufferI、3.2μl四种核苷酸混合液(浓度为2.5mM)、0.8μl CA-1(浓度为20μM)、0.8μl CA-2(浓度为20μM)、2μl上述模板(浓度为20ng/μL)、0.5μl Takara公司的La-taq聚合酶。反应条件为,先94℃ 2分钟;然后94℃ 1分钟,61℃ 1分钟,72℃ 5分钟,共29个循环;最后72℃ 10分钟。
反应结束后,对PCR扩增产物进行1%琼脂糖凝胶电泳检测,PCR扩增出得到2400bp左右正确大小的条带,与预期结果相符。
将上述PCR产物用限制性内切酶Nde I和BamH I进行双酶切后与经同样酶双酶切的质粒pET30(a)(Novagen)进行连接,将连接产物化学转化到大肠杆菌Escherichia coli BL21(DE3)感受态细胞,将转化细胞涂布于添加有50μg/ml卡那霉素的LB平板上筛选阳性克隆,测序结果与野生型头孢菌素酰化酶氨基酸序列相符。
设计引物CA-His-rev:5’-CTCACGCTCGAGAGAAGCACCCGCCGGCACCAGTTCCTGG-3’(其中带下划线碱基代表XhoI位点,其后的9个碱基编码三个连接氨基酸Gly-Ala-Ser)。
提取阳性克隆的质粒作为模板,使用CA-1和CA-His-rev对构建的质粒进行扩增,具体如下:依次向0.5ml薄壁PCR管中加入13.5μl无菌水、25μl 2×GC bufferI、8μl四种核苷酸混合液(浓度为2.5mM)、1μl CA-1(浓度为20μM)、1μlCA-His-rev(浓度为20μM)、1μl模板(浓度为20ng/μL)、0.5μl Takara公司的La-taq聚合酶。按照先94℃ 2分钟;然后94℃ 1分钟,61℃ 1分钟,72℃ 5分钟,共29个循环;最后72℃ 10分钟进行PCR反应。
将得到的PCR扩增产物进行1%琼脂糖凝胶电泳检测,得到2400bp附近的大小的条带,与预期结果相符。
将PCR产物用限制性内切酶Nde I和Xho I进行双酶切后与经同样酶双酶切的质粒pET30(a)进行连接,将连接产物化学转化到大肠杆菌Escherichia coli BL21(DE3)感受态细胞,将转化细胞涂布于添加有50μg/ml卡那霉素的LB平板上筛选阳性克隆。
将阳性克隆接种于含有50μg/ml卡那霉素的LB液体培养基中,37℃ 250rpm过夜培养,提取质粒,送去测序,结果表明,该质粒含有PCR扩增产物,该PCR产物的基因具有序列表中序列2所示的核苷酸,该基因即为野生型头孢菌素酰化酶(CA)的编码基因(GenBank:AAA25690.1),CA的氨基酸序列为序列表中的序列1。
该质粒为将序列表中的序列2插入pET30(a)的中Nde I和Xho I位点间得到的载体,该质粒命名为pET30(a)-CA-His(pET30(a)-CA,示意图如图1)。
也可人工合成序列2作为模板,以CA-1和CA-His-rev作为引物进行PCR扩增,再将该PCR产物经限制性内切酶Nde I和Xho I进行双酶切后与经同样酶双酶切的质粒pET30(a)进行连接,得到pET30(a)-CA-His。
二、构建头孢菌素酰化酶(CA)的突变体CA-113及其突变株E.coli BL21(DE3)/CA-113
思路为从上述步骤一获得的pET30(a)-CA-His出发,利用重叠PCR的技术,在CA野生型的基础上,引入Hisβ57Ala突变。
以上述步骤一获得的pET30(a)-CA-His为模板,首先用如下引物CA-For:
5’-ACAATTCCCCTCTAGAAATAATTTTGTTTAACTTTAAGAAGG-3’(其中下划线代表Xba I内切酶位点)和CA-β57Lower:
5’-GCCGTTATGCGCAAACGCCGGAAAGCCCGGCACGCCCGGCACGGTCAGGCCGATC(其中下划线部分为突变引入位点),进行PCR扩增,退火温度66℃,扩增出突变Hisβ57Ala位点的上游片段;
再同样以pET30(a)-CA-His为模板,使用CA-Rev:
5’-CTCACGCTCGAGAGAAGCACCCGCCGGCACCAGTTCCTGG-3’(其中下划线代表Xho I内切酶识别位点)和CA-β
57Upper:5’-ATGATCGGCCTGACCGTGCCGGGCGTGCCGGGCTTTCCGGCGTTTGCGCATAACGG(其中下划线部分为突变引入位点),进行PCR扩增,退火温度66℃,进行扩增,得到突变Hisβ57Ala位点的下游片段;
最后以上述得到的突变Hisβ57Ala位点的上游片段和突变Hisβ57Ala位点的下游片段为模板,加入CA-For和CA-Rev引物,在退火温度为70℃的条件下进行扩增,得到2400bp附近的PCR产物。
将上述获得的PCR产物用限制性内切酶Xba I和Xho I进行双酶切后,与经同样酶双酶切的质粒pET30(a)进行连接,将连接产物化学转化到大肠杆菌Escherichia coli BL21(DE3)感受态细胞,将转化细胞涂布于添加有50μg/ml卡那霉素的LB平板上筛选阳性克隆。
将阳性克隆接种于含有50μg/ml卡那霉素的LB液体培养基中,37℃ 250rpm过夜培养,提取质粒送去测序,结果表明,该质粒含有PCR扩增产物,该PCR产物的基因具有序列表中序列5所示的核苷酸,该基因命名CA-113,其编码的蛋白命名为CA-113,其氨基酸序列为序列表中的序列3。该质粒为将序列表中的序列5插入pET30(a)的中Xba I和Xho I位点间得到的载体,该质粒命名为pET30(a)-CA-113。
比对野生型CA和突变体CA-113的核苷酸和氨基酸序列如下:
突变体CA-113的核苷酸(序列5)为将野生型CA的核苷酸序列(序列2)的自5’端第886-888位的CAT碱基突变为GCG碱基。
突变体CA-113的氨基酸(序列3)为将野生型CA的核苷酸序列(序列1)的自N末端第296位(β亚基的57位)的组氨酸(H)突变为丙氨酸(A),记为H296A。
也可人工合成序列5作为模板,以CA-1和CA-Rev引物,进行PCR扩增,再将该PCR产物经限制性内切酶Nde I和Xho I进行双酶切后与经同样酶双酶切的质粒pET30(a)进行连接,得到pET30(a)-CA-113。
含有pET30(a)CA-113的菌株命名为E.coli BL21(DE3)/CA-113,即为突变株。
三、利用重叠PCR方法构建头孢菌素酰化酶(CA)的突变体CA-1C及其突变株E.coli BL21(DE3)/CA-1C
思路为从上述步骤二获得的CA-113所得质粒出发,利用重叠PCR的技术,在CA-113的基础上,引入Hisβ70Tyr突变。
以步骤二得到的pET30(a)CA-113为模板,首先用CA-For(序列同步骤二)和CA-β70Lower:5’-AAACGCATAGGTCACGCAATACGCCACTT-3’(其中下划线部分为突变引入位点),进行PCR扩增,退火温度60℃,扩增出突变Hisβ70Tyr位点的上游片段;
再同样以pET30(a)-CA-113为模板,使用CA-Rev(序列同实施例2)和CA-β70Upper:5’-GCGTATTGCGTGACCTATGCGTTTATGGA-3’(其中下划线部分为突变引入位点),退火温度60℃,进行扩增,得到突变Hisβ70Tyr位点的下游片段;
最后以上述得到的突变Hisβ70Tyr位点的上游片段和突变Hisβ70Tyr位点的下游片段为模板,加入CA-For和CA-Rev引物,在退火温度为69℃的条件下进行扩增,得到2400bp左右的PCR产物。
将上述获得的PCR产物用限制性内切酶Xba I和Xho I进行双酶切后,与经同样酶双酶切的质粒pET30(a)进行连接,将连接产物化学转化到大肠杆菌Escherichia coli BL21(DE3)感受态细胞,将转化细胞涂布于添加有50μg/ml卡那霉素的LB平板上筛选阳性克隆。
将阳性克隆接种于含有50μg/ml卡那霉素的LB液体培养基中,37℃ 250rpm过夜培养,提取质粒送去测序,结果表明,该质粒含有PCR扩增产物,该PCR产物的基因具有序列表中序列6所示的核苷酸,该基因命名CA-1C,其编码的蛋白命名为CA-1C,其氨基酸序列为序列表中的序列4。该质粒为将序列表中的序列6插入pET30(a)的中Xba I和Xho I位点间得到的载体,该质粒命名为pET30(a)-CA-1C。
比对野生型CA和突变体CA-1C的核苷酸和氨基酸序列如下:
突变体CA-1C的核苷酸(序列6)为将野生型CA的核苷酸序列(序列2)的自5’端第886-888位的CAT碱基突变为GCG碱基,同时第925-927位的CAT碱基突变为TAT碱基。
突变体CA-1C的氨基酸(序列4)为野生型CA的氨基酸酸序列(序列1)自N末端第296位(β亚基的57位)的组氨酸(H)突变为丙氨酸(A);同时自N末端第309位(β亚基的70位)的组氨酸(H)突变为缬氨酸(Y),记为H296A-H309Y。
也可人工合成序列6作为模板,以CA-1和CA-Rev引物,进行PCR扩增,再将该PCR产物经限制性内切酶Nde I和Xho I进行双酶切后与经同样酶双酶切的质粒pET30(a)进行连接,得到pET30(a)-CA-lC。
含有pET30(a)CA-lC的菌株命名为E.coli BL21(DE3)/CA-1C,即为突变株。
四、头孢菌素酰化酶突变体基因的诱导表达及其纯化
将pET30a转入E.coli BL21(DE3)/pET30a,得到转空载体E.coli BL21(DE3)/pET30a作为空白对照;
将步骤一得到的pET30(a)-CA-His转入E.coli BL21(DE3)/pET30a,得到转空载体E.coli BL21(DE3)/pET30(a)-CA-His作为阳性对照;
分别将上述步骤二和步骤三得到的突变株E.coli BL21(DE3)/CA-113和E.coli BL21(DE3)/CA-1C均在含卡那霉素(50μg/ml)的LB平板上划线,37℃过夜培养,再挑取生长良好的的单菌落接种于10ml LB液体培养基(含50μg/ml卡那霉素)中,同时以E.coli BL21(DE3)/pET30a为空白对照,E.coli BL21(DE3)/pET30(a)-CA-His作为阳性对照,37℃,250rpm过夜培养。
分别将突变株E.coli BL21(DE3)/CA-113、E.coli BL21(DE3)/CA-1C、空白对照和阳性对照,以1∶40的接种量转接于400ml新鲜的液体LB培养基(含50μg/ml卡那霉素)中,继续培养至OD600为0.4。然后加入终浓度为0.5mM的IPTG,在28℃,250rpm条件下培养20h,诱导表达结束后,收集菌体,即为诱导后产物:CA-113、CA-1C、空白对照和阳性对照。
将上述产物分别进行SDS-PAGE检测,结果如图2所示,其中1为阳性对照,2为Marker,3为CA-113,4为CA-1C,5为空白对照,可以看出,CA-1C、CA-113均有57kDa与26kDa的目的条带,而空白对照没有的目的条带,阳性对照有目的条带,说明突变株E.coli BL21(DE3)/CA-113、E.coli BL21(DE3)/CA-1C经诱导后大量表达出头孢菌素酰化酶突变体CA-113和CA-1C。
将上述收集菌体破碎后,分别取细胞上清液10mL,用0.22μm低蛋白质结合的滤膜过滤后,利用蛋白纯化设备( explorerTM station),上样至已结合Ni2+的HiTrapTM Chelating HP柱(购自安玛西亚(Amersham Biosciences),体积1mL),再用咪唑浓度梯度洗脱蛋白,当280nm检测器上出现目标峰时,开始收集至峰消失。分别得到纯化后CA-113(2.81mg/ml)和CA-1C(3.97mg/ml)。
将纯化后CA-113和CA-1C经SDS-PAGE检验,CA-1C、CA-113均有57kDa与26kDa的产物,与目标蛋白野生型头孢菌素酰化酶CA的β亚基和α亚基的理论计算值相近。
采用相同的方法阳性对照E.coli BL21(DE3)/pET30(a)-CA-His的诱导产物进行纯化,得到纯化CA(5.38mg/ml),即为野生型头孢菌素酰化酶。
实施例2、头孢菌素酰化酶突变体的活性检测与转化率
1、头孢菌素酰化酶突变体的活性检测
将由实施例1得到的纯化后的CA-1C和CA-113分别取20μL,与含有3%(质量体积比)的头孢菌素C(石药集团)的pH=8.0,100mM的Tris-HCl的缓冲液180μl混合,得到反应前混合物,在25℃反应l0min后,加入200μl 40%的冰醋酸终止反应,得到反应产物。以纯化CA(野生型头孢菌素酰化酶)为阳性对照,得到阳性对照反应产物。
用高效液相色谱检测反应前混合物和产物,其中用迪马C18液相色谱柱250mm×4.6mm,流动相组成为15%色谱甲醇,7.5%色谱乙腈,1%冰醋酸。以购自Alfa Aesar的7-氨基头孢烷酸作为标准品,以CA-His作为阳性对照(纯化CA,即为野生型头孢菌素酰化酶)。
结果如图3所示,A为反应前混合物(转化初始各组分CPC),B为反应产物(反应结束后各组分),C为标准品7-氨基头孢烷酸,可以看出反应前混合物的保留时间为3.85min;反应产物的保留时间为2.95min。而标准品和阳性对照反应产物的保留时间为2.95;证明反应产物和阳性对照反应产物均为7-氨基头孢烷酸。
在10min内,CA-1C可以产生0.749μmol 7-氨基头孢烷酸,CA-113可以产生0.365μmol 7-氨基头孢烷酸,且即为头孢菌素酰化酶突变体。阳性对照纯化CA(野生型头孢菌素酰化酶)可以产生0.156μmol 7-氨基头孢烷酸。
定义1U为25℃下,在1min内将1μmol底物头孢菌素C转化为产物7-氨基头孢烷酸的活性。计算方法是按照上述活性测定的方法,跟据10min内产生的7-氨基头孢烷酸的总量除以时间。
纯化后CA-1C的活性为943.1U/g(g表示的是纯化后蛋白的质量);
纯化后CA-113的活性为648.0U/g(g表示的是纯化后蛋白的质量);
纯化后CA(野生型头孢菌素酰化酶)的活性为144.0U/g(g表示的是纯化后蛋白的质量)。
2、头孢菌素酰化酶突变体底物转化率
在10ml的反应体系中,按照反应体系3%(质量体积比)投入0.3g底物头孢菌素C钠盐(石药集团),并按每克底物200U共投酶60U;在25℃,在100mM的Tris-HCl缓冲液中,恒pH=8.0的条件下,转化三个小时,所投的酶为突变体纯化后的CA-1C。
通过HPLC检测(检测条件同上)剩余CPC的含量,结果如下:
酶为突变体纯化后CA-1C时,剩余CPC的含量为0.036%,转化率为98%(图4)。而已报道的野生型的头孢菌素酰化酶,在催化底物CPC的反应时,转化率为60%左右(SHIN,Y.C.,J.Y.JEON,et al.(2005).cephalosporin C acylase mutant and method for preparing 7-ACA using same)。
实施例3、头孢菌素酰化酶(CA)的突变体经过氨基酸残基取代的衍生蛋白的构建和活性测定
思路为从上述实施例1中步骤三获得的pET30(a)-CA-1C出发,利用重叠PCR的技术,在CA-1C的基础上,引入Serβ471Ala突变。
以实施例1中的步骤三得到的pET30(a)-CA-1C为模板,首先用CA-For(序列同实施例1,步骤二)和CA-β471Lower:5’-CATAACGAGCCAGCGCGCCATA-3’(其中下划线部分为突变引入位点),进行PCR扩增,退火温度60℃,扩增出突变Serβ471Ala位点的上游片段;
再同样以pET30(a)-CA-1C为模板,使用CA-Rev(序列同实施例1,步骤二)和CA-β471Upper:5’-GGCGCGCTGGCTCGTTATGT 3’(其中下划线部分为突变引入位点),退火温度60℃,进行扩增,得到突变Serβ471Ala位点的下游片段;
最后以上述得到的突变Serβ471Ala位点的上游片段和突变Serβ471Ala位点的下游片段为模板,加入CA-For和CA-Rev引物,在退火温度为69℃的条件下进行扩增,得到2400bp左右的PCR产物。
将上述获得的PCR产物用限制性内切酶Xba I和Xho I进行双酶切后,与经同样酶双酶切的质粒pET30(a)进行连接,将连接产物化学转化到大肠杆菌Escherichia coli BL21(DE3)感受态细胞,将转化细胞涂布于添加有50μg/ml卡那霉素的LB平板上筛选阳性克隆。
将阳性克隆接种于含有50μg/ml卡那霉素的LB液体培养基中,37℃ 250rpm过夜培养,提取质粒送去测序,结果表明,该质粒含有PCR扩增产物,该PCR产物的基因具有序列表中序列7所示的核苷酸,该基因命名CA-5B,其编码的蛋白命名为CA-5B,其氨基酸序列为序列表中的序列8。该质粒为将序列表中的序列7插入pET30(a)的中Xba I和Xho I位点间得到的载体,该质粒命名为pET30(a)-CA-5B。
含有pET30(a)-CA-5B的菌株命名为E.coli BL21(DE3)/CA-5B,即为突变株。
比对野生型CA和衍生蛋白CA-5B的核苷酸和氨基酸序列如下:
衍生蛋白CA-5B的核苷酸(序列7)为将野生型CA的核苷酸序列(序列2)的自5’端第886-888位的CAT碱基突变为GCG碱基,同时第925-927位的CAT碱基突变为TAT碱基,同时第2128-2130位的AGC碱基突变为GCT碱基(即为将突变体CA-1C序列6的自5’端第2128-2130位的AGC碱基突变为GCT碱基)。
衍生蛋白CA-5B的氨基酸(序列8)为野生型CA的氨基酸酸序列(序列1)自N末端第296位(β亚基的57位)的组氨酸(H)突变为丙氨酸(A);同时自N末端第309位(β亚基的70位)的组氨酸(H)突变为缬氨酸(Y),同时自N末端第710位(β亚基的471位)的丝氨酸(S)突变为丙氨酸(A),记为H296A-H309Y-S710A(即为将突变体CA-1C氨基酸序列序列4的自N末端第710位(β亚基的471位)的丝氨酸(S)突变为丙氨酸(A))。
也可人工合成序列7作为模板,以CA-1和CA-Rev引物,进行PCR扩增,再将该PCR产物经限制性内切酶Nde I和Xho I进行双酶切后与经同样酶双酶切的质粒pET30(a)进行连接,得到pET30(a)-CA-5B。
将衍生蛋白表达株E.coli BL21(DE3)/CA-5B在含卡那霉素(50μg/ml)的LB平板上划线,37℃过夜培养,再挑取生长良好的的单菌落接种于10ml LB液体培养基(含50μg/ml卡那霉素)中,同时以E.coli BL21(DE3)/pET30a为空白对照,E.coli BL21(DE3)/pET30(a)-CA-His作为阳性对照,37℃,250rpm过夜培养。
分别将衍生蛋白表达株E.coli BL21(DE3)/CA-5B、空白对照和阳性对照,以1∶40的接种量转接于10ml新鲜的液体LB培养基(含50μg/ml卡那霉素)中,继续培养至OD600为0.4。然后加入终浓度为0.5mM的IPTG,在28℃,250rpm条件下培养5h,诱导表达结束后,8000g离心收集菌体。按照相等的OD重悬后,以衍生蛋白CA-5B、空白对照和阳性对照,分别进行SDS-PAGE检测,可以看出,CA-5B有57kDa与26kDa的目的产物,而空白对照没有的目的条带,阳性对照有目的条带,说明突变株E.coliBL21(DE3)/CA-5B经诱导后大量表达出头孢菌素酰化酶突变体CA-5B。
分别取诱导表达E.coli BL21(DE3)/CA-5B和E.coli BL21(DE3)/pET30(a)-CA-His的菌体破碎的上清液20μL,与含有3%(质量体积比)的头孢菌素C(石药集团)的pH=8.0,100mM的Tris-HCl的缓冲液180μl混合,在25℃反应60min后,加入200μl 40%的冰醋酸终止反应,分别得到反应产物和阳性对照反应产物。
按照实施例2中的液相色谱条件和方法,分析7-氨基头孢烷酸的含量,在保留时间为2.95min处有峰,证明衍生蛋白CA-5B的确产生7-氨基头孢烷酸,且7-氨基头孢烷酸产量(7-氨基头孢烷酸峰的积分面积为948190)为野生型(7-氨基头孢烷酸峰的积分面积为218242)的4.3倍。说明突变体经过氨基酸残基取代的衍生蛋白比野生型的活性提高,其为另一种头孢菌素酰化酶突变体。
头孢菌素酰化酶突变体及其编码基因与应用专利购买费用说明
![]()
Q:办理专利转让的流程及所需资料
A:专利权人变更需要办理著录项目变更手续,有代理机构的,变更手续应当由代理机构办理。
1:专利变更应当使用专利局统一制作的“著录项目变更申报书”提出。
2:按规定缴纳著录项目变更手续费。
3:同时提交相关证明文件原件。
4:专利权转移的,变更后的专利权人委托新专利代理机构的,应当提交变更后的全体专利申请人签字或者盖章的委托书。
Q:专利著录项目变更费用如何缴交
A:(1)直接到国家知识产权局受理大厅收费窗口缴纳,(2)通过代办处缴纳,(3)通过邮局或者银行汇款,更多缴纳方式
Q:专利转让变更,多久能出结果
A:著录项目变更请求书递交后,一般1-2个月左右就会收到通知,国家知识产权局会下达《转让手续合格通知书》。














动态评分
0.0