





专利摘要
葡萄糖剥夺为癌症研究和治疗中令人关注的策略。癌细胞增量调节葡萄糖摄取和代谢,以维持加速的生长和增殖速率。特异性阻断这些过程,可能为葡萄糖转运和代谢在肿瘤发生以及在凋亡中的作用提供新见识。由于实体瘤过大而不适于周围脉管系统,它们遭遇营养物受限供应的微环境,导致在肿瘤的某些区域中形成葡萄糖剥夺的环境。生存在葡萄糖剥夺环境中的癌细胞经受变化以防止葡萄糖剥夺诱导的凋亡。了解癌细胞如何逃避凋亡诱导,亦可能产生有价值的关于如何克服癌细胞中对凋亡诱导的抵抗的信息和知识。本文公开了抑制基础葡萄糖转运、导致肿瘤抑制的新型抗癌化合物,以及用于在动物癌症研究中研究葡萄糖剥夺的新方法。
权利要求
1. 一种式(III)化合物或其盐:
其中,R1选自氢、卤素、烷基、苄基、氨基、硝基、氰基和烷氧基;
其中,R2选自氢、卤素、烷基、苄基、氨基、硝基、氰基和烷氧基;
其中,R3选自氢、卤素、烷基、苄基、氨基、硝基、氰基和烷氧基;
其中,X选自碳、氧、氮和硫;和
其中,Y选自碳、氧、氮和硫。
2. 权利要求1的化合物,
其中,所述化合物选自:
其中,X选自H、3-Cl、3-F、3-CN、4-F、4-CN、4-NO2、4-SO2Me和4,5-Cl2;
其中,Y选自H、2-Cl、2-F、2-OME、2-CH2OH和3-CH2OH;和
其中,Z选自H、2-Cl、2-F、2-OME、2-CH2OH和3-CH2OH。
3. 权利要求1的化合物,
其中,所述化合物选自:
其中,X选自H、3-Cl、3-F、3-CN、4-F、4-CN、4-NO2、4-SO2Me和4,5-Cl2;和
其中,Y选自NHC(O)CH3、NHS(O)2CH3、NHCONH2和CH2OH。
4. 权利要求1的化合物,
其中,R1、R2和R3为氢;和
其中,X和Y为氧。
5. 一种式(IV)化合物或其盐:
其中,R1选自氢、卤素、烷基、苄基、氨基、硝基、氰基和烷氧基;
其中,R2选自氢、烷基、苄基、芳基和杂芳基;
其中,R3选自氢、烷基、苄基、芳基和杂芳基。
6. 权利要求5的化合物,其中,R2为芳基。
7. 权利要求6的化合物,
其中,所述芳基选自2-硝基-5-羟基苯基,2-、3-和4-羟基苯基,2,3-、2,4-、2,5-、2,6-、3,4-和3,5-二羟基苯基,2,3,4-、2,3,5-、2,3,6-和3,4,5-三羟基苯基,2,3,4,5-和2,3,4,6-四羟基苯基,全羟基苯基,2-氨基-5-羟基苯基,2-氰基-5-羟基苯基,2-氟-5-羟基苯基,2-氯-5-羟基苯基,2-溴-5-羟基苯基,2-羧基-5-羟基苯基,2-酮基-5-羟基苯基,2-烷氧基5-羟基苯基和2-烷基-5-羟基苯基。
8. 权利要求5的化合物,其中,R3为芳基。
9. 权利要求6的化合物,
其中,所述芳基选自2-硝基-5-羟基苯基,2-、3-和4-羟基苯基,2,3-、2,4-、2,5-、2,6-、3,4-和3,5-二羟基苯基,2,3,4-、2,3,5-、2,3,6-和3,4,5-三羟基苯基,2,3,4,5-和2,3,4,6-四羟基苯基,全羟基苯基,2-氨基-5-羟基苯基,2-氰基-5-羟基苯基,2-氟-5-羟基苯基,2-氯-5-羟基苯基,2-溴-5-羟基苯基,2-羧基-5-羟基苯基,2-酮基-5-羟基苯基,2-烷氧基5-羟基苯基和2-烷基-5-羟基苯基。
10. 权利要求5的化合物,
其中,R1为氯;和
其中,R2和R3为2-硝基-5-羟基苯基。
11. 一种治疗癌细胞的方法,所述方法包括:
给予需要所述治疗的受试者治疗上有效量的化合物,所述化合物选自式3和4。
12. 权利要求11的方法,其中,所述癌症包括实体恶性瘤。
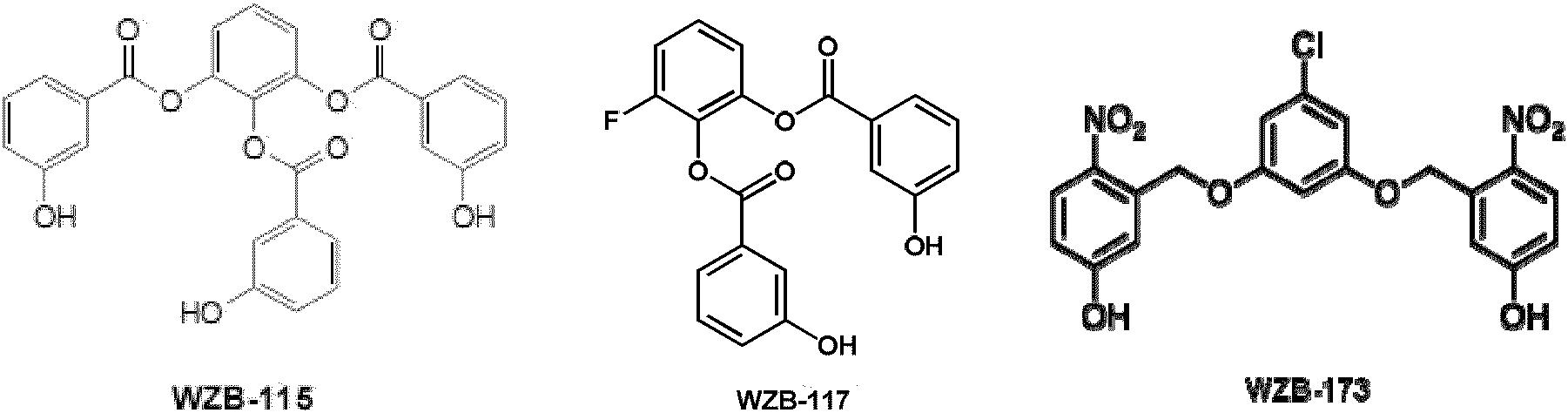
13. 权利要求11的方法,其中,所述癌症增量调节基础葡萄糖转运。
14. 权利要求11的方法,
其中,所述给予包括以下方法之一:经口、局部、动脉内、胸膜内、鞘内、心室内、皮下、腹膜内、静脉内、囊内和卡氮芥糯米纸胶囊剂。
15. 权利要求11的方法,其中,所述受试者为人。
16. 权利要求11的方法,其进一步包括给予第二种癌症药物。
17. 权利要求16的方法,
其中,所述第二种癌症药物选自甲氨蝶呤、盐酸阿霉素、氟尿嘧啶、依维莫司、咪喹莫特、阿地白介素、阿仑单抗、培美曲塞二钠、盐酸帕洛司琼、苯丁酸氮芥、氨基乙酰丙酸、阿那曲唑、阿瑞吡坦、依西美坦、奈拉滨、三氧化二砷、奥法木单抗、贝伐珠单抗、阿扎胞苷、盐酸苯达莫司汀、贝沙罗汀、博来霉素、硼替佐米、卡巴他赛、盐酸依立替康、卡培他滨、卡铂、盐酸柔红霉素、西妥昔单抗、顺铂、环磷酰胺、氯法拉滨、异环磷酰胺、阿糖胞苷、达卡巴嗪、地西他滨、达沙替尼、地加瑞克、地尼白介素-毒素连接物、狄诺塞麦、盐酸右雷佐生、多西他赛、拉布立酶、盐酸表柔比星、奥沙利铂、艾曲波帕乙醇胺、甲磺酸艾日布林、盐酸厄洛替尼、磷酸依托泊苷、盐酸雷洛昔芬、托瑞米芬、氟维司群、来曲唑、非格司亭、磷酸氟达拉滨、普拉曲沙、吉非替尼、盐酸吉西他滨、吉西他滨-顺铂、吉妥珠单抗奥佐米星、甲磺酸伊马替尼、曲妥珠单抗、盐酸拓扑替康、替伊莫单抗、罗咪酯肽、伊沙匹隆、帕利夫明、二甲苯磺酸拉帕替尼、来那度胺、亚叶酸钙、醋酸亮丙瑞林、脂质体盐酸丙卡巴肼、替莫唑胺、普乐沙福、acetidine、甲苯磺酸索拉非尼、尼洛替尼、枸橼酸他莫昔芬、罗米司亭、紫杉醇、盐酸帕唑帕尼、培门冬酶、泼尼松、盐酸丙卡巴肼、proleukin、利妥昔单抗、罗米地辛、滑石粉、甲苯磺酸索拉非尼、苹果酸舒尼替尼、沙利度胺、坦罗莫司、托瑞米芬、曲妥珠单抗、帕尼单抗、硫酸长春碱、长春新碱、伏林司他、唑来膦酸及其任何组合。
说明书
相关申请
本申请要求保护2010年3月24日提交的美国临时申请号61/317,062的权益,所述申请的全部内容通过引用结合到本文中。
关于联邦赞助研究的声明
美国国家科学基金通过创新奖助金(HER-0227907)的合作,部分地赞助本工作。美国政府可在本发明中具有一定权利。
发明背景
自2008年起,癌症已超过心血管疾病,成为美国的第一杀手,仅在2008年,估计有565,650个美国人死于癌症。已提出关于癌症的起因的不同理论,并且已制定和开发许多策略用于对抗所述疾病。一些癌症例如乳腺癌的死亡率在过去30年间已显著减少,这主要是因为早期检测而不是因为治疗,而其它癌症例如肺癌和胰腺癌的死亡率实际上却增加。绝对和迫切需要新方法用于进一步改进现存癌症疗法以及用于治疗对其而言仍无有效疗法的那些癌症。葡萄糖剥夺可具有成为这类新型且有效抗癌策略之一的潜力,这是因为在理解Warburg效应上取得的最新进展所致,Warburg效应即癌细胞对增加的葡萄糖转运和葡萄糖代谢(主要是糖酵解)增加的和“沉迷的”依赖。
几乎所有癌症的共同特征之一以及亦可能是它们的共同弱点之一,为增加的葡萄糖摄取以及对于将葡萄糖作为细胞生长和增殖的结构单元来源、能源或二者的增加的依赖性。尽管癌症并非单一疾病,但是不同的癌症特别是实体恶性瘤的确共享一些共同特征。这类共同特征之一为,它们均比正常细胞长得快并因此需要更多合成前体和更多能量来维持其加速的生长和增殖速率。正常细胞可将不同的化学品(例如氨基酸、脂质和葡萄糖)用作其能源。
与典型细胞相比,用于癌细胞的生物合成材料和能量的优选来源为葡萄糖。例如,健康结肠细胞从短链脂肪酸特别是丁酸盐中得到其能量供应的60-70%。丁酸盐经由单羧酸转运体MCT1转运穿过结肠上皮的管腔膜。已发现显示降低水平的MCT1的癌样品,表达高亲和力葡萄糖转运体GLUT1,这表明在转变成恶性期间就结肠上皮中的能源/生物合成来源而言存在从丁酸盐到葡萄糖的转换。
与体内正常细胞相比,几乎所有癌细胞在体内具有增加的葡萄糖供应和代谢,这是已通过正电子发射断层扫描(PET)提供的最强有力的证据片段(图14)。在癌症的PET扫描中,将作为不可代谢葡萄糖类似物的18F标记的2-脱氧葡萄糖(2-DG或FDG)用作示踪物。在扫描中发亮的区为捕获较多FDG的器官、组织、细胞以及癌症。斑点越亮表示FDG浓度越高。与许多其它PET扫描一样,此特异性PET扫描显示,原发癌和转移癌(接近肺和腋窝)二者均比周围正常细胞含有更高的FDG浓度,提供了癌细胞相对于正常细胞具有增加的葡萄糖摄取的有力证据。对不同癌症类型(包括原发癌和继发转移癌二者)的PET扫描显示,与环绕肿瘤的正常细胞和组织相比,几乎所有研究的肿瘤“捕获”明显更多FDG。此外,PET扫描研究已将预后不良和增加的肿瘤侵占性与增加的葡萄糖摄取和增量调节的葡萄糖转运体一致地关联。尽管已提出多种理论来解释在癌细胞内使用葡萄糖的机制,但是本领域存在几乎一致共识:不论葡萄糖在其吸收后如何被癌细胞使用,在几乎所有恶性瘤中葡萄糖摄取为增加的。癌细胞增加的葡萄糖摄取及其伴随增加的葡萄糖代谢可以是、应当是并且已成为用于深入细致的的基础和临床研究以及用于开发新抗癌疗法的一般靶标。
在1920年代,Warburg发现,即使在充足氧气存在下,癌细胞仍优选通过在胞质溶胶中的糖酵解来代谢葡萄糖,而不像在正常细胞中通过在线粒体中的氧化磷酸化来代谢葡萄糖。这看似矛盾,因为糖酵解在生成ATP上效率更低。已表明这类向糖酵解的转换赋予癌细胞一些选择优势,用于在独特的肿瘤微环境中生存和增殖。因加速的生长速率和不足的供氧所致,结节中的大部分癌细胞处于含氧量低的环境,迫使癌细胞通过增加葡萄糖转运体、糖酵解酶和线粒体代谢抑制剂的表达来向糖酵解转变。然而,Warburg效应不能仅通过对缺氧的适应来解释,因为即使在存在充足氧气时,癌细胞仍优选糖酵解。这有可能涉及其他的分子机制。
近期研究已显示,在Warburg效应中观察到的现象亦可在癌基因激活中发现,所述现象为增加的葡萄糖消耗和减少的氧化磷酸化,以及伴随的大幅增加的乳酸盐产生。发现Ras在突变时促进糖酵解。发现Akt的激活增加糖酵解的速率,这部分地因为其通过HIFα促进糖酵解酶表达的能力。将此推测为促成癌细胞的高糖酵解性质的主要因素。亦发现Myc (原癌基因和转录因子)增量调节多个代谢基因的表达。还发现肿瘤抑制基因例如p53涉及代谢调节。所有这些近期研究结果表明,癌细胞中的Warburg效应不只是仅在糖酵解中独立变化的结果,更是通过多个信号转导途径中已知和未知的串扰网络形成的广泛交流的生物学后果。这些途径涉及细胞生长、增殖以及线粒体代谢和葡萄糖代谢二者,所述代谢响应在供氧和营养物供应上的变化。理解Warburg效应中的这类广泛的信号转导网络,对于理解和对抗癌症而言为必需的。
一些最近的研究已集中在糖酵解酶特别是丙酮酸激酶(PK)上。这些研究已显示,癌细胞中增加的葡萄糖转运和糖酵解似乎指向癌细胞中结构单元的产生(大分子的生物合成),并且为细胞分裂和增殖做准备,而非作为提供生物能(ATP)的方式。尽管通常认为有氧糖酵解是癌症的代谢标志,但是其与肿瘤发生的因果关系仍不清楚。糖酵解基因包括癌细胞中最多增量调节的基因集之一。在肿瘤中显著增量调节的基因中有PK,其调节糖酵解的最终限速步骤。在哺乳动物中存在PK的4个同种型:L和R同种型在肝和红细胞中表达;M1同种型在大多数成体组织中表达;和M2同种型为在胚胎发育期间表达的M1的剪接变体。值得注意的是,已报道肿瘤组织唯一地表达丙酮酸激酶的胚胎M2同种型。由于其几乎普遍存在于癌细胞中,因此将PKM2称为肿瘤特异的,并且目前将其在人血浆中的存在用作诊断多种癌症的分子标志。正常增殖细胞和肿瘤细胞二者均表达PKM2。PKM2调节葡萄糖碳的比例,所述葡萄糖碳被引入合成过程(无活性二聚体形式)或用于糖酵解能量产生(高活性四聚体形式,糖酵解酶复合体的组分)。在癌细胞中,二聚体形式的PKM2总是占优势的。四聚体和二聚体形式的PKM2之间的转换允许肿瘤细胞在具有不同供氧和营养物供应的环境中生存。两种形式之间的转换在肿瘤细胞中调节糖酵解通量。这些研究结果表明,PKM2为代谢传感器,其以葡萄糖供应依赖性方式调节细胞增殖、细胞生长和凋亡性细胞死亡。发现PKM2的核易位足以诱导非胱天蛋白酶依赖性细胞死亡和同种型特异性细胞死亡。这些结果显示,肿瘤标记物PKM2在肿瘤细胞的非胱天蛋白酶依赖性细胞死亡中起广泛作用,并因此将此糖酵解酶定义为用于癌症疗法开发的新靶标。
两项近期研究证实,PKM2通过与磷酸酪氨酸基序结合来调节,导致促进增加的细胞生长和肿瘤发展。PKM2增强糖酵解中间体用于大分子生物合成和肿瘤生长的应用。这些研究结果说明此代谢表型在癌细胞生长中的独特优势。似乎PKM2的表达以及从氧化磷酸化向有氧糖酵解的转换对于维持癌生长和增值而言为绝对需要的。因此,抑制糖酵解以及PKM2可形成新且有效的抗癌策略。这些新研究结果意义重大,因为它们几乎完全改变了我们对Warburg效应在癌症中的生物学功能的常规理解,曾认为所述效应用于在含氧量低的条件下生物合成ATP。
葡萄糖为大部分细胞中代谢的必需底物。由于葡萄糖为极性分子,因此通过生物膜转运需要特定的转运蛋白。通过肠上皮细胞和肾上皮细胞的顶膜转运葡萄糖依赖于次级主动Na+/葡萄糖同向转运体SGLT-1和SGLT-2的存在,所述SGLT-1和SGLT-2利用由Na+离子顺其电化学梯度向下的协同转运提供的能量在细胞内浓缩葡萄糖。通过细胞膜促进扩散葡萄糖由葡萄糖载体(对于溶质载体家族2,蛋白质符号GLUT,基因符号SLC2)另外催化,所述葡萄糖载体属于转运促进剂超家族(主要的促进剂超家族),所述超家族包括有机阴离子和阳离子转运体、酵母己糖转运体、植物己糖/质子同向转运体以及细菌糖/质子同向转运体。通过这类转运体的分子移动通过促进的扩散来进行。此特征使得这些转运体为非能量依赖性的,不同于常常需要ATP的存在以驱动其易位机制并且在ATP/ADP比率降至过低时停止的主动转运体。
基础葡萄糖转运体(GLUT)充当葡萄糖通道并且是维持细胞的基本葡萄糖需求所必需的。这些GLUT在细胞中组成地表达和有功能,并且不受胰岛素调节(或者对胰岛素不敏感)。所有细胞在线粒体中利用糖酵解和氧化磷酸化二者,但在氧充足时极大地依赖于氧化磷酸化,在氧剥夺(缺氧)时则转换成糖酵解,如在癌症中发生的转换。在糖酵解中,葡萄糖转变为丙酮酸盐并且在所述过程中生成2个ATP分子(图15)。癌细胞因其较快的增殖速率所致,主要处于含氧量低的(低氧)状态。因此,癌细胞利用糖酵解(乳酸形成)作为其优势的葡萄糖代谢途径。这类糖酵解转换不仅使癌症具有更高的转移和侵袭潜力,而且还增加癌症对于在糖酵解方面的外部干扰的易损性,这是因为癌细胞“沉迷”于葡萄糖和糖酵解。基础葡萄糖转运的减少可能限制对癌细胞的葡萄糖供应,导致迫使癌细胞减缓生长或饿死的葡萄糖剥夺。Thompson小组发现激活的Akt在成胶质细胞瘤细胞系中导致受激的需氧葡萄糖代谢,并发现所述细胞随后在撤去葡萄糖时死亡。这为癌细胞对葡萄糖浓度变化非常敏感以及葡萄糖剥夺可在癌细胞中诱导死亡提供直接证据。
在正常细胞中,如图15所示,靶细胞通过一个或多个基础葡萄糖转运体(GLUT)摄取胞外葡萄糖。细胞使用的GLUT取决于细胞类型和生理需要。例如,GLUT1负责所有细胞类型中低水平的基础葡萄糖转运。所有GLUT蛋白包含12个跨膜结构域并且通过促进扩散(非能量依赖性过程)来转运葡萄糖。GLUT1可能通过改变其构象来转运葡萄糖进入细胞。根据此模型,GLUT1向细胞外或细胞内暴露单一的底物结合部位。葡萄糖与一个部位的结合触发构象改变,将葡萄糖释放到膜的另一侧。转基因和敲除动物研究的结果支持这些转运体在控制葡萄糖利用、葡萄糖储存和葡萄糖传感方面的重要作用。GLUT蛋白在其动力学上不同并且针对其服务的细胞类型的需要而定制。尽管特定的细胞类型可表达多于一种GLUT蛋白,但是癌症经常过表达GLUT1 (其为高亲和力葡萄糖转运体,并且其表达水平与癌症的侵袭力和转移潜能相关),表明葡萄糖转运的增量调节在癌细胞生长和癌症恶性的严重性上的重要性。亦发现GLUT1表达显著高于任何其他葡萄糖转运体的表达。在一项研究中,测试的全部23种肿瘤为GLUT1阳性并且GLUT1为主要表达的葡萄糖转运体。此外,FDG摄取和GLTU1表达二者似乎都与增加的肿瘤大小有关。在数种肿瘤(包括NSCLC、结肠癌、膀胱癌、乳腺癌和甲状腺癌)中,增加的GLUT1表达不仅赋予恶性表型,还预示着较低的总体存活率。基于所有的这些观察结果,可以想象的是,通过基础葡萄糖转运抑制来抑制癌症生长,可以是阻止癌症生长以及改善预后和存活时间的有效方式。
证据指出,与正常细胞相比,癌细胞对葡萄糖剥夺更敏感。许多研究强烈表明,基础葡萄糖转运抑制诱导凋亡并阻止癌细胞生长。首先,已显示抗血管发生是限制癌症生长并引起癌症消融的非常有效的方式。本质上,所述抗血管发生方法将减少新血管形成并且在肿瘤结节之内和周围达到血管标准化。这严重限制了到达癌细胞的对肿瘤生长所需的营养物。葡萄糖为通过抗血管发生剥夺的关键营养物之一。在这种意义上,基础葡萄糖转运的抑制在限制对癌细胞的营养物供应上可看作抗血管发生疗法的备选方法。因此,抗血管发生策略的成功间接支持将限制对癌细胞的葡萄糖供应作为相关但新型策略的潜在功效。其次,涉及糖酵解的多种酶的抑制剂已用于抑制糖酵解过程中的不同步骤,并且已显示具有显著的抗癌功效。已靶定的糖酵解酶包括:己糖激酶,催化糖酵解第一步的酶;ATP柠檬酸裂解酶;以及更近的丙酮酸脱氢酶激酶(PDK)。在测试的糖酵解抑制剂中,发现3-溴丙酮酸和己糖激酶抑制剂完全根除经治疗小鼠中的晚期糖酵解肿瘤。靶定线粒体糖酵解酶乳酸脱氢酶A (LDH-A)的化合物在体外和体内均显示显著的抗癌活性。此结果表明线粒体功能和胞质糖酵解之间的强烈联系。用于PET扫描以定位转移的示踪物2-DG,已用作抗癌临床试验中的葡萄糖竞争剂和糖酵解抑制剂。这些研究以及其他相关研究亦已显示,这些抑制剂在癌细胞中诱导凋亡作为癌细胞杀伤机制。可从所有这些公布的研究中得出两个重要结论。(1)抑制糖酵解多个步骤的化合物在体外和体内均减少癌细胞生长,和(2)在癌细胞中抑制糖酵解的多个步骤之一诱导凋亡并且为有效的抗癌策略。其亦强烈表明,抑制葡萄糖转运(临糖酵解前的步骤以及糖酵解和细胞内所有葡萄糖代谢的第一限速步骤)将对癌细胞产生类似于糖酵解抑制或者可能比糖酵解抑制更严重的生物学后果。此外,葡萄糖转运可能是与下游糖酵解靶标相比更好的靶标,因为1)已知葡萄糖转运体在癌细胞中高度增量调节,2)通过在第一步限制葡萄糖供应以及因此造成绝对的胞内葡萄糖缺乏,将防止任何潜在的胞内葡萄糖相关的补偿/补救途径,癌细胞可利用所述补偿/补救途径自我救护和避免细胞死亡。
为于使抑制基础葡萄糖转运成为成功的抗癌策略,其必须在不显著伤害正常细胞的情况下杀死癌细胞。一些实验观察结果表明,事实的确如此。由于癌细胞偏爱将葡萄糖用作能源以及糖酵解在癌细胞中增量调节,因此抑制糖酵解的化合物可杀死癌细胞同时不伤害正常细胞,所述正常细胞可将脂肪酸和氨基酸用作备选能源。
最近报道,向多种肺癌和乳腺癌细胞系中添加抗GLUT1抗体显著降低癌细胞的葡萄糖摄取速率和增殖,导致凋亡的诱导。此外,所述抗体增强诸如顺铂、紫杉醇和吉非替尼等癌症药的抗癌作用。这些结果明确指出,抑制GLUT1介导的葡萄糖转运的物质,在单独作用时或在与其他抗癌治疗药联用时,对抑制癌细胞生长以及在癌细胞中诱导凋亡有效。两个最近的出版物进一步支持这些研究结果,在所述出版物中,发现葡萄糖转运抑制剂fasentin使癌细胞对经历由抗癌药顺铂或紫杉醇诱导的凋亡敏感,以及发现抗癌化合物芹菜素在mRNA和蛋白质水平上减量调节GLUT1。提出将GLUT1的减量调节作为芹菜素可能的抗癌机制。所有这些新研究结果指明以下方向,葡萄糖转运抑制剂可能敏化并协同其他抗癌药,以进一步增强药物的抗癌功效。本文公开了在抑制基础葡萄糖转运和诱导凋亡上有效性为fasentin或芹菜素有效性的2-5倍的化合物和方法。
在使用葡萄糖剥夺的一项近期研究中,发现在高浓度生长因子中生长的细胞在撤去生长因子后显示出对细胞死亡增加的易感性。此易感性与生长因子撤去之后在糖酵解速率上的变化幅度相关。为了研究糖酵解产物的可用性变化是否影响线粒体开始的凋亡,通过操纵细胞培养基中的葡萄糖水平来人为地限制糖酵解。与生长因子撤去相同,葡萄糖限制导致Bax易位、线粒体膜电位下降以及细胞色素c释放至胞质溶胶。相比之下,通过过表达GLUT1来增加细胞自主的葡萄糖摄取,在生长因子撤去之后显著延迟凋亡。这些结果表明,生长因子的主要功能为调节葡萄糖摄取和代谢,并因此维持线粒体内环境稳定以及使细胞生长所需的合成代谢途径能够进行。亦发现涉及葡萄糖摄取和糖酵解定型的三个基因(GLUT1、己糖激酶2和磷酸果糖激酶1)的表达,在生长因子撤去后迅速降至几乎不可检测的水平。所有这些研究表明,葡萄糖剥夺已是用于研究癌症的非常有价值且频繁使用的方法。亦可通过抑制基础葡萄糖转运来造成胞内葡萄糖剥夺。因自细胞培养基中去除葡萄糖所致的葡萄糖剥夺和因抑制葡萄糖转运/葡萄糖代谢所致的葡萄糖剥夺之间的区别为,葡萄糖去除首先产生缺乏葡萄糖的胞外环境,而葡萄糖转运/葡萄糖代谢的抑制在不改变或甚至增加胞外葡萄糖浓度的情况下,产生缺乏葡萄糖的胞内环境。葡萄糖转运抑制剂的使用应当能够补充和替代传统的葡萄糖剥夺。此外,通过降低胞外葡萄糖浓度的传统葡萄糖剥夺不能用于动物,而葡萄糖转运的抑制剂可用于动物,这创建了在体内研究癌症和治疗癌症的新方法。
发明简述
本文公开了式(I)化合物或其盐,其中R1选自氢、烷基、苄基、芳基和杂芳基部分;R2选自氢、烷基、苄基、芳基、杂芳基和荧光标签;R3选自氢、卤素、烷基、苄基、芳基、杂芳基、氨基、氰基和烷氧基。在一些实施方案中,如本领域技术人员所公认的,两个R1基团可独立选择,且因此不同。在其他实施方案中,当R1基团不同时,可将R1表示为R1’和R1’’以表明R1部分之间的不同。
在一些实施方案中,可进一步限定式(I)化合物包括以下种类,其中R1为芳基官能团,选自2-、3-和4-羟基苯基,2,3-、2,4-、2,5-、2,6-、3,4-和3,5-二羟基苯基,2,3,4-、2,3,5-、2,3,6-和3,4,5-三羟基苯基,2,3,4,5-和2,3,4,6-四羟基苯基以及全羟基苯基。在其他实施方案中,可进一步限定式(I)化合物,其中R2为荧光标签,选自香豆素、丹酰、罗丹明、荧光素和羧基萘并荧光素。在一些实施方案中,式(I)化合物由以下分子组成,其中R1和R2为3-羟基苯基,R3为氢原子。
本文公开了式(II)化合物或其盐,其中R1选自氢、烷基、苄基、芳基和杂芳基;R2选自氢、烷基、苄基、芳基和杂芳基;X选自氢、卤素、烷基、苄基、芳基、杂芳基、氨基、氰基和烷氧基;Y选自氢、卤素、烷基、苄基、芳基、杂芳基、氨基、氰基和烷氧基。
在一些实施方案中,可进一步限定式(II)化合物包括以下种类,其中R1为芳基官能团,选自2-、3-和4-羟基苯基,2,3-、2,4-、2,5-、2,6-、3,4-和3,5-二羟基苯基,2,3,4-、2,3,5-、2,3,6-和3,4,5-三羟基苯基,2,3,4,5-和2,3,4,6-四羟基苯基以及全羟基苯基。在其他实施方案中,式(II)化合物由以下分子组成,其中R1和R2为3-羟基苯基,X为氟,Y为氢。
本文公开了一系列式(III)化合物或其盐,其中R1选自氢、卤素、烷基、苄基、氨基、硝基、氰基和烷氧基;R2选自氢、卤素、烷基、苄基、氨基、硝基、氰基和烷氧基;R3选自氢、卤素、烷基、苄基、氨基、硝基、氰基和烷氧基;X选自碳、氧、氮和硫;Y选自碳、氧、氮和硫。
在一些实施方案中,式(III)化合物可选自下列化合物;
。
在一些实施方案中,式(III)化合物可选自下列化合物;
,并且
其中X选自H、3-Cl、3-F、3-CN、4-F、4-CN、4-NO2、4-SO2Me和4,5-Cl2。在其它实施方案中,式(III)化合物由以下分子组成,其中R1、R2和R3为氢,X和Y为氧。
本文公开了式(IV)化合物或其盐,其中R1选自氢、卤素、烷基、苄基、氨基、硝基、氰基和烷氧基;R2选自氢、烷基、苄基、芳基和杂芳基;R3选自氢、烷基、苄基、芳基和杂芳基。在一些实施方案中,式(IV)化合物是这样的分子,其中R1为氯,R2和R3为2-硝基-5-羟基苯基。
本文公开了用于治疗癌症的方法,其包括给予需要这类治疗的受试者治疗上有效量的选自式1、2、3和4的化合物。
在一些实施方案中,所述癌症为实体恶性瘤,其通过在称为Warburg效应的过程中从氧化磷酸化向糖酵解的生物学转变,增量调节基础葡萄糖转运。在一些实施方案中,可通过选自以下的任何方法给予人受试者所述化合物:经口、局部、动脉内、胸膜内、鞘内、心室内、皮下、腹膜内、静脉内、囊内和卡氮芥糯米纸胶囊剂。
在一些实施方案中,可将来自式1、2、3和4的化合物与一种或多种化学治疗剂组合给予人受试者或患者,作为增强一种或多种所述治疗上有用化合物的功效的方法。在其它实施方案中,来自式1、2、3和4的化合物可与之组合给予的化学治疗剂选自:甲氨蝶呤、盐酸阿霉素、氟尿嘧啶、依维莫司、咪喹莫特、阿地白介素、阿仑单抗、培美曲塞二钠、盐酸帕洛司琼、苯丁酸氮芥、氨基乙酰丙酸、阿那曲唑、阿瑞吡坦、依西美坦、奈拉滨、三氧化二砷、奥法木单抗、贝伐珠单抗、阿扎胞苷、盐酸苯达莫司汀、贝沙罗汀、博来霉素、硼替佐米、卡巴他赛(cabazitaxel)、盐酸依立替康、卡培他滨、卡铂、盐酸柔红霉素、西妥昔单抗、顺铂、环磷酰胺、氯法拉滨、异环磷酰胺、阿糖胞苷、达卡巴嗪、地西他滨、达沙替尼、地加瑞克、地尼白介素-毒素连接物(denileukin difitox)、狄诺塞麦(denosumab)、盐酸右雷佐生、多西他赛、拉布立酶、盐酸表柔比星、奥沙利铂、艾曲波帕乙醇胺(eltrombopaq olamine)、甲磺酸艾日布林(eribulin mesylate)、盐酸厄洛替尼、磷酸依托泊苷、盐酸雷洛昔芬、托瑞米芬、氟维司群、来曲唑、非格司亭、磷酸氟达拉滨、普拉曲沙、吉非替尼、盐酸吉西他滨、吉西他滨(gemcitibine)-顺铂、吉妥珠单抗奥佐米星、甲磺酸伊马替尼、曲妥珠单抗(trastuzamab)、盐酸拓扑替康、替伊莫单抗、罗咪酯肽(romadepsin)、伊沙匹隆、帕利夫明、二甲苯磺酸拉帕替尼、来那度胺、亚叶酸钙、醋酸亮丙瑞林、脂质体盐酸丙卡巴肼、替莫唑胺、普乐沙福、acetidine、甲苯磺酸索拉非尼、尼洛替尼、枸橼酸他莫昔芬、罗米司亭(romiplostim)、紫杉醇、盐酸帕唑帕尼(pazopanib hydrochloride)、培门冬酶、泼尼松、盐酸丙卡巴肼、proleukin、利妥昔单抗、罗米地辛、滑石粉(Talc)、甲苯磺酸索拉非尼(sorafenic tosylate)、苹果酸舒尼替尼、沙利度胺、坦罗莫司、托瑞米芬、曲妥珠单抗(trastuzumub)、帕尼单抗(pantiumumab)、硫酸长春碱、长春新碱、伏林司他和唑来膦酸。
另外的特征和优势将部分地在下列描述中阐述,以及部分地从所述描述中显而易见,或者可通过本发明的实践而知悉。本发明的目标和优势将通过在随附权利要求中明确指出的要素和组合来实现和达到。
要理解的是,上述概述和下列详述二者仅为示例性和解释性的,并且不限制所要求保护的本发明。
附图简述
结合到本说明书中并构成本说明书一部分的附图,说明了本发明的一些实施方案,并连同描述,用来解释本发明的原理。
图1显示初始葡萄糖转运的抑制剂1a和2a。
图2显示可能来自1p、9a和2a的水解产物的葡萄糖摄取结果。这些可能的水解产物未显示显著的基础葡萄糖摄取抑制,表明所述抑制是因原始化合物而非水解产物所致。
图3显示α-PGG和β-PGG的分子结构。PGG具有葡萄糖核,其通过在葡萄糖的羟基和没食子酸之间形成的酯键,与五个没食子酰基连接。α-PGG和β-PGG为结构异构体。α-PGG及其衍生物为亲水的并且可能胞外地作用于细胞膜蛋白。
图4显示基础葡萄糖转运的新抑制剂WZB-25、WZB-26和WZB-27的结构。
图5显示α-PGG处理在癌细胞中诱导细胞死亡,并且所述细胞死亡主要通过凋亡介导。图A显示,如通过细胞生存力(MTT)测定所测定,α-PGG处理导致细胞生存力减少70-80%。图B显示,如通过凋亡(ELISA)测定所测定,α-PGG处理在HeLa细胞中导致凋亡大于2倍的增加。图C显示,如通过抗体偶联的流式细胞术研究所测定,HeLa细胞的α-PGG处理导致G1期细胞减少而凋亡细胞显著增加(约3倍)。
图6显示如通过蛋白质印迹分析所测定的p53激活和失活。当用25μM α-PGG处理HeLa、RKO或MCF-7细胞时,发现p53蛋白在HeLa细胞中未被激活,但在RKO细胞为已激活的。
图7显示α-PGG及其衍生物在HeLa、RKO和MCF-7癌细胞中抑制葡萄糖摄取。在用3H标记的2-DG处理之前,将细胞用α-PGG处理20分钟。在加入2-DG 30分钟之后,将细胞收集、裂解并对其各自的葡萄糖摄取计数。样品:1.模拟的;2.胰岛素(100nM);3. α-PGG (30μM);4. WZB-25 (30μM);5. WZB-26 (30μM);和6. WZB-27 (30μM)。
图8A-C显示α-PGG及其衍生物以剂量依赖性方式在HeLa细胞中抑制基础葡萄糖转运。
图9显示通过α-PGG或α-PGG衍生物诱导的葡萄糖转运抑制的时程。除在不同的时间(从1、5、10直至30分钟)终止葡萄糖摄取之外,用与前述相同的方式进行葡萄糖摄取测定。
图10显示α-PGG诱导Akt,而α-PGG衍生物不诱导Akt。用α-PGG或其衍生物处理过表达胰岛素受体的CHO细胞。处理之后,裂解细胞并通过对磷酸化Akt特异的抗体来分析蛋白质。
图11显示,与在其正常细胞对应物相比,PGG衍生的化合物在癌细胞中诱导更多的细胞死亡。左图显示,使用25mM化合物处理人肺癌细胞(H1299)或正常肺细胞(NL20)。在处理后48小时,进行细胞生存力测定以确定细胞杀伤的百分比。将未经化合物处理的细胞用作对照(100%基线)。右图显示用于处理人乳腺癌细胞(MCF-7)和正常乳腺细胞(MCF-12A)的化合物。在与左图肺癌细胞所用条件相同的条件下,进行细胞生存力测定。
图12显示细胞生存力测定和凋亡测定。图A显示在RKO结肠癌细胞中的细胞生存力测定。RKO细胞含有高水平的p53,而RKO E6细胞含有低得多水平的p53。图B显示H1299肺癌细胞中的凋亡测定,其使用经切割的PARP蛋白(89 kDa)作为胱天蛋白酶3的指示剂,因为PARP是已激活胱天蛋白酶3的底物。
图13显示葡萄糖转运的示意图。
图14显示原发性和转移性人癌症的PET扫描。
图15显示α-PGG在3T3-L1脂肪细胞中的胰岛素样活性。A.如通过葡萄糖摄取测定所测定的与胰岛素的葡萄糖转运刺激活性相比的α-PGG的葡萄糖转运刺激活性。B. α-PGG诱导胰岛素样GLUT4膜易位,如在经不同物质的细胞诱导之后,使用GLUT4特异性抗体通过荧光共聚焦显微术所示。α-PGG诱导胰岛素样GLUT4易位,如在环绕细胞的环状结构中。
图16显示新型PGG衍生物。
图17显示两个试验化合物抑制剂的结构。化合物WZB-27和WZB-115为衍生自PGG的多酚化合物。与PGG不同,它们不具有胰岛素样葡萄糖摄取刺激活性。相反的是,它们仅具有有效的葡萄糖转运抑制活性和抗癌活性,如在葡萄糖摄取测定和MTT细胞生存力测定中所证实。
图18显示细胞生存力测定和凋亡测定(W25 = WZB-25,W27 = WZB-27)。A. RKO结肠癌细胞中的细胞生存力测定。RKO细胞含有高水平的p53,而RKO E6细胞含有低得多水平的p53。B. α-PGG在RKO中比在RKO-E6细胞中诱导更强的生存力降低效应。RKO细胞具有较高水平的p53,而RKO-E6细胞为p53缺陷型。***p < 0.001,**p < 0.01。C. A549肺癌细胞中的凋亡测定,其使用经切割的PARP蛋白(89 kDa)作为胱天蛋白酶3的指示剂,因为PARP是已激活胱天蛋白酶3的底物。低葡萄糖(正常的5%)处理的样品作为阳性对照。
图19显示,与其非癌对应物相比,癌细胞表达更多的GLUT1蛋白以及在葡萄糖摄取上受化合物抑制更大。使用或不使用化合物处理癌细胞及其非癌对应物,然后测定其各自的葡萄糖摄取。A.使用或未使用WZB-27处理的H1299肺癌细胞及其非癌NL20细胞的葡萄糖摄取测定。B. MCF7癌细胞及其非癌MCF12A细胞的葡萄糖摄取测定。C.癌细胞和非癌细胞中GLUT1蛋白表达的蛋白质印迹分析,其使用对GLUT1 (H43片段)特异的抗体。β-肌动蛋白用作蛋白质上样对照。
图20显示化合物注射后的血糖水平。将化合物W27 (= WZB-27)或W115 (= WZB-115) IP注入禁食的Balb/c健康小鼠中,并在注射后多次测定血糖水平。N = 5只/组。PBS + DMSO组为溶媒对照。
图21显示葡萄糖抑制剂和抗癌药的组合进一步降低癌细胞生存力。在不存在或存在10 μM WZB-115或30μM WZB-27的情况下,使用抗癌药顺铂(2.5 μM用于W27 (= WZB-27)研究或5 μM用于W115 (=WZB-115)研究)或泰素(2.5 μM)处理H1299、A549肺癌细胞或MCF7乳腺癌细胞。通过MTT测定法测定细胞生存力。所述化合物的存在显著增加通过顺铂或泰素诱导的癌细胞死亡。将此实验重复三次并将结果表示为平均值±标准差。
图22显示在癌细胞的葡萄糖摄取抑制和细胞生存力上,内部化合物抑制剂与fasentin之间的比较研究。在葡萄糖摄取测定和细胞生存力(MTT)测定二者中,将已知的基础葡萄糖化合物fasentin和内部制备的化合物抑制剂并列比较。将模拟处理样品的葡萄糖摄取或细胞生存力任意指定值为100%。A. H1299癌细胞中的葡萄糖摄取测定。对于所有化合物浓度均为30 μM。B.三种不同癌细胞系中的细胞生存力测定。对于所有化合物浓度均为60 μM。
图23显示化合物WZB-115在59个癌细胞系中的NCI抗癌活性筛选结果。将WZB-115送至NCI,使用包括9种癌症类型的59个癌细胞系进行抗癌活性筛选。以单一浓度10 μM完成试验。将模拟处理癌细胞的生长速率用作基线100%。小于100%的任何生长速率表示抑制。因其有前景的抗癌活性谱所致,NCI推荐使用五个不同的浓度再次测试所述化合物,以确定其在这些癌细胞系中的IC50。
图24显示用于鉴定改进的基础葡萄糖转运抑制剂的一般方案。
图25显示用于结构修饰的数个先导化合物和区域。
图26显示束缚类似物(tether analog)的初始集合。
图27显示所提议键合类似物的最小能量结构。
图28显示酚基的生物异构体类似物。
图29显示核心芳环的初始集合。
图30显示移植到裸小鼠上的人肺癌A549的肿瘤大小比较。在治疗后8周获取照片。使用雄性NU/J裸小鼠(7-8周大),购自Jackson实验室(Bar Harbor, Maine)并且提供来自Harlan实验室(Indianapolis, Indiana)的已辐照Teklad Global 19%蛋白质啮齿类动物食物。为了确定化合物WZB-117对人NSCLC肿瘤异种移植物生长的体内功效,收集指数生长的A549细胞,并用PBS重悬浮以达到5×106个细胞/25μl混悬液的终浓度。将25μl细胞悬液皮下注入每只小鼠的右胁。此时,将小鼠随机分成两组:用PBS/DMSO (1:1,v/v)治疗的对照组(n=10),以及用WZB-117 (15mg/kg)治疗的WZB-117治疗组(n=10)。将化合物WZB-117溶于PBS/DMSO (1:1, v/v)。自肿瘤细胞接种之日起,每天用PBS/DMSO混合物或化合物WZB-117 (15mg/kg)给予小鼠腹膜内注射。A.肿瘤生长曲线。动物肿瘤研究表明,通过每天以10 mg/kg体重的剂量注射WZB-117,经化合物治疗肿瘤的肿瘤大小平均比经模拟治疗小鼠的肿瘤大小小约75%。B.小鼠肿瘤照片。左边的荷瘤小鼠为经模拟治疗的,而右边的小鼠是用WZB-117治疗的。C.小鼠体重测定。D.经WZB-117治疗小鼠的体重组成与经模拟治疗小鼠的体重组成的比较。
图31显示WZB-117通过抑制Glut1来抑制葡萄糖转运。A.和B. WZB-117以剂量依赖性方式在红细胞(RBC)中抑制葡萄糖转运。C.和D. WZB-117以剂量依赖性方式在RBC衍生的“内翻外”囊泡(IOV)中抑制葡萄糖转运。E. WZB-117在RBC衍生的“右翻外”囊泡(ROV)中抑制葡萄糖转运。
图32显示癌细胞的WZB-117治疗诱导ER应激、凋亡以及在糖酵解酶上的改变。A. WZB-117治疗以类似于葡萄糖剥夺的方式增量调节ER应激蛋白BiP。B. WZB-117治疗诱导PARP的切割,表明凋亡诱导由p53介导。C. WZB-117治疗以类似于葡萄糖剥夺的方式在癌细胞中增量调节关键的糖酵解酶PKM2。
图33显示使用或未使用WZB-117治疗的小鼠的摄食。在研究期间,PBS/DMSO治疗组和WZB-117治疗组之间没有摄食的改变。自肿瘤细胞接种起,每7天测定各组的摄食。
图34显示使用或未使用WZB-117治疗的荷瘤裸小鼠的血糖测定。通过血糖监测器测定各小鼠的血糖水平,并且在临IP注射WZB-117 (15mg/kg)之前和注射后每30分钟测定血糖水平。测定期间小鼠可得到食物。以平均值±标准差表达数据。在紧接化合物注射之后或动物研究期间或动物研究之后,未发现未治疗组和化合物WZB-117注射组之间在血糖水平上的显著差异。
图35显示化合物WZB-117杀死的癌细胞显著多于杀死的非癌细胞。使用或不使用WZB-117将A. A549肺癌细胞和B. MCF7乳腺癌细胞处理48小时,然后用MTT测定法测定其各自的生存率。经模拟处理的细胞作为对照(100%生存力)用于比较。以相同的方式处理非癌NL20和MCF12A细胞以用于比较。
图36显示如通过克隆形成测定所证实的WZB-117的抗癌活性。使用WZB-117或较弱的抑制剂WZB-134处理或者不使用化合物(模拟的)将生长在培养皿中的三个癌细胞系A549、H1299 (肺癌)和MCF7 (乳腺癌)处理48小时。然后使经处理的细胞回到无化合物的正常细胞培养基中生长2周,随后用结晶紫(crustal violet)染色并对存活克隆的数量计数。染色斑(克隆)越少且越小,抑制越高。
图37显示数个新型葡萄糖转运抑制剂WZB-115、117和173的结构。化合物117为115的类似物,而173为醚键类似物。化合物WZB-117和WBZ-173衍生自化合物WZB-115,所述WZB-115为用于我们早前癌症研究的多酚模型化合物。WZB-115衍生自称为五没食子酰基葡萄糖(PGG)的天然抗癌抗糖尿病化合物。WZB-117和WZB-173在结构上非常类似于115,但是与WZB-115相比,二者在结构上简化且在功能上优化。因此,WZB-117和WZB-173在其抗癌活性上比WZB-115更有效,并且在溶液和细胞培养基中在结构上亦比115更稳定。
发明详述
现在将通过参考一些更详细的实施方案并偶尔参考附图来描述本发明。然而,这些发明可以不同的形式实施,并且不应解释为限于本文所述实施方案。但是,提供这些实施方案以使本公开内容透彻和完整,并且将向本领域技术人员充分地传达本发明的范围。
除非另有定义,否则本文所用的所有技术和科学术语的意义,与这些发明所属领域的普通技术人员通常理解的意义相同。本文用于描述本发明的术语仅用于描述具体的实施方案,且不意图其为本发明的限制。除非文段另有明确指示,否则在本发明的描述和随附权利要求中所用的单数形式“一”、“一个”和“所述”,亦意图包含复数形式。本文提及的所有出版物、专利申请、专利和其他参考文献,均通过引用以其整体结合。
除非另有指示,否则用于说明书和权利要求中表达组分的量、反应条件等的所有数字,应理解为在所有情况中受术语“约”修饰。因此,除非表示反对,否则下列说明书和所附权利要求中陈述的数值参数为近似值,其可根据本发明试图获得的所需特性而改变。丝毫不试图限制权利要求范围的等同物的原则的应用,应根据有效位数和普通四舍五入法来解释各数值参数。
虽然描述本发明广泛范围的数值范围和数值参数为近似值,但是尽可能精确地记录具体实施例中陈述的数值。然而,因在其各自测试法中存在的标准差所致,任何数值固有地必然含有一定的误差。在本说明书各处给出的各数值范围包含各更窄的数值范围(其落入所述更宽的数值范围内),犹如所述更窄的数值范围均在本文中明确写出。
此外,当以Markush群组或其他备选分组的方式描述本发明的特征或方面时,本领域技术人员将认识的是,亦因此以Markush群组或其他群组成员的任何独立成员或子群的方式描述本发明。
定义
“烷基”应指仅由元素氢和碳组成的任何化合物,其中原子唯一地通过单键连接在一起。术语烷基亦可扩展为意指仅由元素氢、氟和碳组成的任何化合物,其中原子唯一地通过单键连接在一起。这组氟化化合物亦可称为“氟代烷”、“氟烷基”、“氟烷基基团”和“碳氟化合物”。烃的实例包括但不限于甲基、乙基、正丙基、异丙基、环丙基、正丁基、仲丁基、异丁基、叔丁基、环丁基、正戊基、异戊基、新戊基、环戊基、正己基、环己基、叔己基、正庚基、正辛基、正癸基和金刚烷基。氟化化合物的实例包括但不限于氟甲基、二氟甲基、三氟甲基、1,1,1-三氟乙基、2,2-二氟乙基和全氟乙基。
“苄基”应用来描述这样的取代基或分子片段,其具有与有机化合物的RC6H4CH2-相关的结构。经取代的苄基化合物亦可描述为与亚甲基(-CH2-)亚单元连接的任何芳基或杂芳基环系。苄基的实例包括但不限于苄基;2、3和4-卤代苄基;2、3和4-烷基苄基;2、3和4-氰基苄基;2、3和4-酮基苄基;2、3和4-羧基苄基;2、3和4-氨基苄基;2、3和4-硝基苄基;2、3和4-羟基苄基;2、3和4-烷氧基苄基;二取代苄基和三取代苄基衍生物。
“芳基”应意指化合物上衍生自简单芳环的任何官能团。实例包括但不限于苯基;2-、3-和4-羟基苯基;2,3-、2,4-、2,5-、2,6-、3,4-和3,5-二羟基苯基;2,3,4-、2,3,5-、2,3,6-和3,4,5-三羟基苯基;2,3,4,5-和2,3,4,6-四羟基苯基;全羟基苯基;2、3和4-卤代苯基;2、3和4-烷基苯基;2、3和4-氰基苯基;2、3和4-酮基苯基;2、3和4-羧基苯基;2、3和4-氨基苯基;2、3和4-硝基苯基;2、3和4-羟基苯基;2、3和4-烷氧基苯基;二取代苯基和三取代苯基衍生物。
“杂芳基”应意指化合物上衍生自杂芳环的任何官能团。杂芳物质包含杂原子或者除氢或碳之外的原子,包括氧、氮、硫、磷、硅和硼。实例包括但不限于呋喃、苯并呋喃、噻吩、苯并噻吩、吡咯、吲哚和硼杂苯。
“卤代”和“卤素”应指周期表第17族的任何元素,由氟、氯、溴、碘和砹组成。
“胺”和“氨基”应指含有碱性氮的任何有机化合物或官能团,所述碱性氮带有孤对电子。胺衍生自氨,并且可为伯胺、仲胺和叔胺的。胺的实例包括但不限于氨、甲胺、二甲胺、三甲胺、乙胺、二乙胺、三乙胺、乙基二甲胺、异丙胺、二异丙胺、二异丙基乙胺、二苯胺、二苄胺、叔丁胺、苯胺(analine)和吡啶。
认为“氰基”应与有机官能团腈同义,其包含与氮原子三键连接的碳。氰化物在自然中倾向于为高毒性的,并且通常作为盐发现。
“烷氧基”应意指与氧单键连接的烷基。烷氧基的范围很大,其范围从甲氧基到许多芳基烷氧基。烷氧基的实例包括但不限于甲氧基、乙氧基、正丙氧基、异丙氧基、正丁氧基、叔丁氧基和苯氧基。
“荧光标签”、“荧光分子”、“荧光团”和“荧光标记”应意指以下分子的任何部分,科学家或研究员已将所述分子化学地连接,以帮助检测与其连接的分子。荧光标签的实例包括但不限于香豆素、丹酰、罗丹明、荧光素、羧基萘并荧光素和荧光蛋白。
“盐”应指由来自式1、2、3和4的化合物之一的阴离子衍生物与阳离子物质配对形成的离子物质。阳离子物质可包括但不限于锂、钠、钾、铷、铯、铍、镁、钙、锶、钡、铝、铜、锌、铁、铬、锰、镍、钯、铂、铟、铑和砷。
“治疗上有效的”,当用于描述在方法中施用的化合物的量时,是指化合物达到所需生物效应的量,例如,导致基础葡萄糖转运抑制的量。
“抑制”或“终止”应意指减少、抑制、损伤、消除、杀死或其组合。
文段中“降低”基础葡萄糖转运应意指降低癌细胞内葡萄糖转运的效率。
α-PGG衍生的第1代化合物的结构;WZB-25、WZB-26和WZB-27
用于合成化合物WZB-25、WZB-26和WZB-27的方法如下:
室温下,向1,6-脱水-β-D-葡萄糖(100 mg,0.62 mmol)的无水乙腈(25 mL)溶液中加入酰氯2 (863 mg,1.88 mmol)。室温搅拌30 min后,向反应混合物中加入DMAP (241 mg,1.97 mmol),将混合物搅拌24h并去除溶剂,通过色谱法在硅胶上纯化粗制品,以83%产率得到730 mg 3。1H NMR (CDC13) δ 7.60-7.31 (m,51H),5.88 (s,1H),5.69 (t,1H,J= 3.0 Hz),5.29 (d,3H,J= 5.3 Hz),5.23 (s,4H),5.11-5.02 (m,13H),4.98 (d,1H,J= 6.2 Hz),4.28 (d,1H,J= 7.4 Hz),3.97 (q,1H,J= 6.2,7.4 HZ)。
向3 (420 mg,0.29 mmol)的无水THF (20.0 mL)溶液中加入10%披钯碳(21 mg,0.02 mmol),室温下将混合物在氢气气氛下搅拌过夜。将混合物滤过硅藻土(Celite),用甲醇和二氯甲烷稀释滤液,并滤过硅藻土三次直至溶液澄清。去除溶剂,以64%产率得到粗制品4。
室温下,向3-甲氧基儿茶酚(140 mg,1.00 mmol)的无水乙腈(10 mL)溶液中加入酰氯2 (941 mg,2.05 mmol)。室温搅拌30 min后,向反应混合物中加入DMAP (268 mg,2.20 mmol),将混合物搅拌2天并去除溶剂,通过色谱法在硅胶(25 % EA/己烷)上纯化粗制品,以72%产率得到708 mg 6。1H NMR (CDC13) δ 7.64 (d,4H,J= 16.7 Hz),7.51-7.33 (m,31H),7.16 (d,1H,J= 8.2 Hz),7.06 (d,1H,J= 8.4 Hz),5.18 (d,4H,J= 7.0 Hz),5.06 (d,8H,J= 11 Hz),3.96 (s,3H);13C NMR (CDC13) δ 164.0、163.6、153.1、152.8、152.3、144.1、143.2、137.6、137.5、136.5、136.4、132.3、128.6、128.4、128.3、128.2、128.1、128.0、127.8、127.7、126.6、123.9、123.8、115.4、110.2、109.7、109.6、75.3、71.2、56.4。
向6 (500 mg,0.51 mmol)的无水THF (20.0 mL)溶液中加入10%披钯碳(43 mg,0.04 mmol),室温下将混合物在氢气气氛下搅拌12h。然后将混合物滤过硅藻土,通过色谱法在硅胶上浓缩并纯化滤液,得到9.5 mg 7。大部分化合物7在硅胶上分解。1H NMR (CDC13) δ 8.30 (brs,6H),7.31-7.13 (m,5H),7.04 (d,1H,J= 8.1 Hz),6.95 (d,1H,J= 8.3 Hz),3.83 (s,3H)。
室温下,向连苯三酚(126 mg,1.00 mmol)的无水乙腈(15 mL)溶液中加入酰氯2 (1.40 g,3.05 mmol)。室温搅拌30 min后,向反应混合物中加入DMAP (391 mg,3.20 mmol),将混合物搅拌24h并去除溶剂,通过色谱法在硅胶(25 % EA/己烷作为洗脱液)上纯化粗制品,以41%产率得到570 mg 9。1H NMR (CDC13) δ 7.53 (s,4H),7.49-7.20 (m,50H),5.10 (s,4H),5.01 (s,8H),4.94 (s,2H),4.85 (s,4H);13C NMR (CDC13) δ 163.6、163.0、152.7、144.2、143.6、143.3、137.4、137.3、136.3、136.1、135.2、128.6、128.5、128.4、128.2、128.1、128.0、127.9、127.8、126.3、123.6、123.1、120.9、109.6、75.2、75.1、71.2。
向6 (260 mg,0.29 mmol)的无水THF (15.0 mL)溶液中加入10%披钯碳(16 mg,0.024 mmol),室温下将混合物在氢气气氛下搅拌过夜。然后将混合物滤过硅藻土,通过色谱法在硅胶(25% EA/己烷)上浓缩并纯化滤液,得到11.3 mg 10。化合物10在柱上分解。1H NMR (CDC13) δ 8.24 (brs,4H),7.47-7.41 (m,1H),7.34-7.32 (m,2H),7.17 (s,4H),7.09 (s,2H),2.92 (brs,5H)。
衍生自α-PPG的第1代化合物的评价
最初将化合物1a和2a制备为α-PGG的潜在抗糖尿病药类似物(图1)。考虑到这类化合物紧密的SAR,猜测更刚性的支架(即苯环)可能增强活性。令人惊讶且意外地发现,这两种化合物在30 μM的浓度下对宫颈癌细胞(HeLa)、结肠癌细胞(RKO)和乳腺癌细胞(MCF-7)抑制基础葡萄糖转运(表1),而非具有胰岛素样活性。
表1.1a和2a在不同癌细胞系中的葡萄糖转运抑制活性(%)
与未经化合物处理的细胞对照(视为0%抑制)相比,如通过标准葡萄糖摄取测定法所测定,化合物1a和2a亦在H1299细胞中抑制基础葡萄糖转运分别达58.4 ± 6.3%和86.1 ± 1.0% (表2)。在H1299细胞中使用MTT细胞增殖测定法测试,发现其对癌细胞生长的抑制活性分别为36.0 ± 6.1%和39.9 ± 5.0% (未经化合物处理的细胞对照视为0%抑制)。
考虑到葡萄糖转运的抑制对于开发新抗癌剂的潜在效用,研究了这些化合物作为葡萄糖转运抑制剂和癌细胞增殖抑制剂二者的构效关系。基于这两种化合物制备了许多衍生物,以了解在中心芳环上对三羟基苯基酯的需要以及对这三种酯的需要。
通过用一组经取代的苯甲酰基卤化物将一系列二羟基苯和三羟基苯酰化,制备了所需类似物。选择一组一羟基苯甲酰基卤化物、二羟基苯甲酰基卤化物和三羟基苯甲酰基卤化物以及甲氧基苯甲酰基卤化物作为酰化剂。羟基苯甲酰基卤化物的合成在方案1中概述。将可市购的酚3a-f全苄化并将所得的酯水解,将酸转化成酰氯5a和苄氧基苯甲酰氯5b、5c、5d、5e和5f。必需的甲氧基取代苯甲酰基卤化物制备自可市购的羧酸。
表2.制备的化合物,其在H1299肺癌细胞中在基础葡萄糖转运和细胞生长上诱导的抑制活性
方案1.苄氧基苯甲酰氯5的合成
根据核心芳环,选择连苯三酚8 (用2a表示)和3-甲氧基儿茶酚6a (X = OMe,Y = H,用1a表示)。选择两个经卤素取代的酚6b (X = H,Y = Cl)和6c (X = F,Y = H)来提供p-供体(类似于甲氧基)而不是吸电子基团。未取代的儿茶酚6d (X,Y = H)包含在内。如在方案2中所示,然后使这些酚的每一种与每种酰氯5a-f以及数个甲氧基取代苯甲酰氯偶联。偶联之后,通过催化氢化将苄氧基酯脱保护。总产率在表2中显示。
方案2.类似物1、2、7和9的合成
如表2所示,使用标准葡萄糖摄取测定法测试化合物。简单说来,在葡萄糖摄取测定法之前,使用或不使用化合物(30 μM)一式三份将H1299癌细胞处理10 min。在不存在或存在化合物的情况下,通过在无葡萄糖KRP缓冲液中将细胞与0.2 Ci/mL [3H]2-脱氧葡萄糖(比活,40 Ci/mmol)一起孵育30 min,来测定细胞葡萄糖摄取。在用冰冷的PBS洗涤细胞并通过0.2 N NaOH裂解细胞之后,将细胞裂解物转移至闪烁计数瓶并通过液体闪烁计数来定量细胞裂解物中的放射性。利用MTT测定法一式六份(hexad)在每孔接种有5,000个细胞的96孔组织培养板中进行细胞生长测定。在存在或不存在所述化合物(30 μM)的情况下将细胞孵育48 h。孵育之后,使用96孔SPECTRAMAX?吸光度/荧光板读数器(Molecular Devices)测定细胞生存力。
在一些实施方案中,将化合物1a和2a与其中核心芳环经氟或氯取代的衍生物(1b和1c)进行比较,显示这两种化合物在葡萄糖转运抑制测定中具有类似于化合物2a且显著优于1a的活性。显然核心芳环上的卤素取代很重要。未取代的核心芳环1d显示低于1a和2a的抑制水平。为了确定酚羟基的必要性,我们制备了1a和2a的数个衍生物,其中用甲氧基取代OH基(9a–d,7a–d)。这些化合物一致显示更低水平的葡萄糖转运抑制。仅化合物9b和9d (2,6-二甲氧基苯甲酰基和3-甲氧基苯甲酰基)显示适度水平的抑制。
在一些实施方案中,系统地去除酚羟基,制备一系列二羟基和一羟基衍生物。3,5-二羟基和3,4-二羟基衍生物总体上显示良好水平的葡萄糖转运抑制。在两个系列中,三苯甲酰基衍生物(2b和2c)以及氟取代和氯取代衍生物显示>90%抑制,而甲氧基取代的衍生物(1e和1h)显示几乎没有至没有抑制。显示>90%葡萄糖转运抑制的所有化合物亦显示癌细胞生长速率降低约40%。
在一些实施方案中,去除额外的羟基提供在2-、3-和4-位的一组一羟基化合物。2-羟基系列一致显示较弱的葡萄糖转运抑制。4-羟基系列尽管显示较弱的葡萄糖转运抑制,但的确提供具有葡萄糖转运抑制>85%的化合物。这些化合物显示癌细胞生长速率降低>35%。3-羟基系列显示优秀的葡萄糖转运抑制,三苯甲酰基衍生物2f显示葡萄糖转运抑制>99%。此化合物亦显示最高水平的细胞生长抑制,约为60%。通过分析表2中的所有数据,得到线性相关系数R = 0.817 (R2 = 0.667),表明约2/3 (66.7%)的癌细胞生长抑制活性来自基础葡萄糖转运的抑制活性。
一般来说,侧臂苯甲酰基的3-位上羟基的存在,对于葡萄糖转运抑制和癌细胞生长抑制二者而言均为重要的。此3-羟基可与氯取代或氟取代的核心苯环连接或者为三苯甲酰基体系的部分。未取代的或供电子的取代基导致葡萄糖转运活性抑制的显著减小。
在其他实施方案中,酚酯的水解产物(8、6c、10、11和12)可表现出葡萄糖摄取抑制活性。如图2所示,这些化合物均不具有任何葡萄糖摄取抑制活性。类似地,这些化合物均未显示在抗癌筛选上的任何活性。总的来说,已合成多酚酯化合物的文库作为基础葡萄糖转运的新型抑制剂和新型抗癌剂,其具有潜在的新靶标——基础葡萄糖转运。
通过SAR研究鉴定出有效且具选择性的基础葡萄糖转运抑制剂
在构效关系(SAR)研究期间(其中已合成超过80个PGG类似物并通过葡萄糖摄取测定和功能测定进行分析),亦鉴定出葡萄糖摄取的一组抑制剂,其具有与PGG相对的活性。偶然发现的是,这些抑制剂引起癌细胞生长抑制和癌细胞死亡。利用这些新研究结果,决定化学合成更有效且更具选择性的抑制剂,并将它们用于癌症研究。
考虑到PGG类似物初始集合的紧密SAR,猜测没食子酰基附于其上的更刚性的支架,可能提供增强的效力和选择性。制备了PGG类化合物的数个新类似物。两种化合物基于芳香核(与葡萄糖核相对)。对所有化合物的假设为,刚性中心核将防止与葡萄糖核相关的任何构象可动性。当分析这些化合物(WZB-26和WZB-27)时,发现其具有基础葡萄糖转运抑制活性而不具有任何胰岛素样活性。因此,这些化合物为动物细胞的基础葡萄糖转运的选择性抑制剂。当使用这些化合物处理不同的癌细胞系时,发现它们亦在宫颈癌细胞(HeLa)、结肠癌细胞(RKO)和乳腺癌细胞(MCF-7)中抑制基础葡萄糖转运。此结果表明,这些化合物为广泛的基础葡萄糖转运抑制剂;抑制测试的全部三种癌细胞系。基于这两种化合物制备了许多衍生物,以了解在中心芳环上对三羟基苯基酯的需要以及对这三种酯的需要(图16)。简单说来,使用受保护的羟基苯甲酸酰化一系列二或三羟基苯。偶联之后,去除保护基(苄基)以提供靶化合物——三苯甲酰基衍生物(3)或苯基取代的二苯甲酰基衍生物(5)。
化合物WZB-26和WZB-27为首先制备的化合物。两种化合物均显示类似的基础葡萄糖转运抑制(约85%)。最初,中心芳环上的取代模式令人关注。因为WZB-27具有三个没食子酰基,而WZB-26仅具有2个没食子酰基,但是具有额外的甲氧基。完成了多种化合物的制备,以探测中心芳环上的其他取代基将对抑制水平的影响。在一些实施方案中,可将制备的化合物命名为WZB-89 (F取代)、WZB-90 (Cl取代)和WZB-110 (H取代)。WZB-89和WZB-90二者均显示与WZB-26和-27类似的活性水平,而未取代的WZB-101显示显著降低的活性水平。显然,如在WZB-26、-27、-89和-90中所示,在3-或4-位中一些类型的π-供电子团有益于活性。WZB-26和WZB-27二者均具有三个酚羟基。为了确定这些羟基是充当H键供体还是H键受体,制备了4种类似物,其中用OMe基团取代OH基团(WZB-76、-81、-101、-102)。这些化合物的活性显著降低,明确表明对于H键供体(即酚OH)的需要。接着,通过系统去除羟基来检测对于没食子酰基(即3,4,5-三羟基苯甲酰基)的需要。制备了一系列二羟基(3,5-二羟基和3,4-二羟基)和一羟基(2-OH、3-OH和4-OH)衍生物。在所有情况下,不论侧臂苯甲酰酯上羟基的数量或位置,中心芳环上具有甲氧基或无取代的类似物均提供显著更低水平的抑制。制备的类似物中,3,5-二羟基(WZB-111、WZB-113、WZB-114)、3,4-二羟基(WZB-119、WZB-121、WZB-112)和3-一羟基(WZB-115、WZB-117、WZB-118)衍生物显示95-99%的抑制水平。鉴于在制药工业存在对于开发更简单、更低分子量抑制剂的关注,因此首先将化合物WZB-27和WZB-115用于生物研究,并且在后续合成和测定中将使用更佳的化合物。表3表明,以制备对基础葡萄糖转运更有效和更具选择性的抑制剂为目标,我们已系统地合成超过100种不同的化合物来研究所述化合物的SAR。如表3中所示,这些抑制剂中的某些是迄今为止报道的最佳基础葡萄糖转运抑制剂并且可用于未来的临床研究。
表3.制备的类似物以及基础葡萄糖转运抑制
α-PGG在人结肠癌细胞、宫颈癌细胞和乳腺癌细胞中诱导凋亡
当使用α-PGG处理RKO(结肠)、HeLa(宫颈)和MCF-7(乳腺)人癌细胞时,发现所述处理导致明显的细胞死亡(图5A),且所述细胞死亡主要地(如果不是唯一地)由凋亡引起(图5B & 5C)。α-PGG在正常(非癌)细胞中不会引起大量凋亡(未显示数据),表明所述化合物更多地对癌细胞显示增加的细胞毒性。
与其正常对应物相比,PGG-衍生的化合物优先在癌细胞中诱导细胞死亡
癌细胞强烈地依赖葡萄糖作为其优选的能源,并且已提出将葡萄糖剥夺作为抗癌策略。为了使这些化合物成为有效的抗癌剂,它们必须能够杀死多于正常细胞的癌细胞。细胞杀伤(或细胞生存力)测定显示,这些化合物中的某些特别是WZB-27 (= W27),与其非癌细胞对应物(NL20或MCF12A细胞,表3)相比,优先杀死癌细胞(NSCLC H1299和乳腺癌MCF7)。这些结果表明,这些化合物具有成为抗癌剂的优秀潜力。基于此观察结果,推测可确定最佳的化合物和药物浓度,其将最小地影响正常细胞,同时对癌细胞造成最大的损伤。
使用WZB-27、WZB-115以及两种已知的抗癌药(顺铂和泰素)完成比较测定。在表4中,显示了癌细胞系以及正常细胞系中的细胞死亡百分比。化合物WZB-27和WZB-115杀死与泰素几乎相同百分比的肺癌细胞系H1299,同时杀死显著更少的正常肺细胞系NL20。比较乳腺癌细胞系MCF7时,WZB-27和WZB-115相对于泰素杀死稍少的细胞,但与顺铂相比杀死更多的细胞。WZB-27和WZB-115二者与泰素或顺铂相比,均杀死更少的正常乳腺细胞系MCF12A。WZB-27与泰素或顺铂相比,杀死更少的正常乳腺细胞系MCF12A,而WZB-115与顺铂或泰素相比,不会杀死更多的正常细胞。表4所示结果表明,化合物在癌细胞中的细胞毒性与顺铂和/或泰素相当或更好,同时它们与抗癌药相比,在非癌(“正常”)细胞中表现出更小的细胞毒性。
表4.癌细胞与正常细胞中通过化合物诱导的%细胞死亡的比较a,b,c
癌细胞系过表达GLUT1以及与其非癌细胞对应物相比,化合物在癌细胞中抑制更多的葡萄糖摄取
为了确定与其非癌细胞对应物相比,在癌细胞中杀伤增加的可能原因,进行比较研究以确定化合物处理对葡萄糖摄取的作用。发现与在非癌细胞系NL20或MCF12A中的降低相比,化合物WZB-27在癌细胞系H1299和MCF7中在葡萄糖摄取上产生更大的降低(图19A和19B)。蛋白质印迹分析显示,这些相同的癌细胞系与其非癌对应物相比,表达显著更高水平的GLUT1蛋白(图19C)。在癌细胞系中观察到的葡萄糖摄取的更大降低,与这些细胞中更高的GLUT1水平有关。
α-PGG在RKO(结肠)细胞中激活p53,但在HeLa(宫颈)或MCF-7(乳腺)癌细胞中不激活p53
发现α-PGG在三个癌细胞系中诱导凋亡之后,研究了凋亡机制的知识(即,凋亡是否与p53状态有关)。使用抗p53抗体的蛋白质印迹分析显示,α-PGG在RKO细胞中导致p53的激活,但在HeLa或MCF-7细胞中不导致p53的激活(图6)。此结果表明,在HeLa和MCF-7细胞中诱导的凋亡为非p53依赖性的。此结果既令人关注又重要,因为其显示α-PGG可利用非p53依赖性机制在某些癌细胞中诱导凋亡,非p53依赖性机制应在超过50%的所有人类癌症中有效诱导凋亡,在所述人类癌症中,p53已突变且为无功能的。
此结果表明,与在RKO细胞中不同,α-PGG在HeLa细胞和MCF-7中诱导的凋亡不通过p53或p53信号转导途径介导。因此,α-PGG在HeLa细胞中诱导的凋亡为非p53依赖性的。
PGG及其衍生物在人癌细胞系中抑制基础葡萄糖转运
将基础葡萄糖转运的抑制推测为α-PGG诱导的癌细胞死亡的原因。合成PGG衍生物并且在不同的人癌细胞系中与α-PGG一起测试。发现这些衍生物在宫颈癌细胞系、结肠癌细胞系和乳腺癌细胞系中抑制基础葡萄糖转运(图7)。
胰岛素不影响葡萄糖摄取,表明测定的葡萄糖为基础葡萄糖转运,而非胰岛素介导的葡萄糖转运。基于此观察结果,我们进一步猜测,显著的葡萄糖转运抑制可能是凋亡(特别是在HeLa细胞中)的原因。
α-PGG及其衍生化合物以剂量依赖性方式在HeLa细胞中抑制基础葡萄糖转运
使用不同浓度的α-PGG或其衍生化合物WZB-25和WZB-27,以确定基础葡萄糖转运的抑制是否为剂量依赖性的。实验结果表明,α-PGG及其衍生物以剂量依赖性方式抑制基础葡萄糖转运,并且在抑制转运上,α-PGG似乎比其衍生物稍更有效(图8)。亦发现30 μM任何化合物在HeLa细胞中导致基础葡萄糖转运的约50%抑制(图8),表明全部4种化合物在HeLa细胞中抑制基础葡萄糖转运上几乎同样有效。此结果使我们得到结论,我们可使用α-PGG或用其衍生物代替
用于葡萄糖转运抑制的组合物和方法专利购买费用说明
![]()
Q:办理专利转让的流程及所需资料
A:专利权人变更需要办理著录项目变更手续,有代理机构的,变更手续应当由代理机构办理。
1:专利变更应当使用专利局统一制作的“著录项目变更申报书”提出。
2:按规定缴纳著录项目变更手续费。
3:同时提交相关证明文件原件。
4:专利权转移的,变更后的专利权人委托新专利代理机构的,应当提交变更后的全体专利申请人签字或者盖章的委托书。
Q:专利著录项目变更费用如何缴交
A:(1)直接到国家知识产权局受理大厅收费窗口缴纳,(2)通过代办处缴纳,(3)通过邮局或者银行汇款,更多缴纳方式
Q:专利转让变更,多久能出结果
A:著录项目变更请求书递交后,一般1-2个月左右就会收到通知,国家知识产权局会下达《转让手续合格通知书》。







动态评分
0.0