

IPC分类号 : C12N9/90,C12N15/61,C12N5/10,C12P33/00





专利摘要
本发明涉及一种雷公藤三萜合酶TwOSC3蛋白及其编码基因,将Twosc3基因的cDNA克隆到真核表达载体pYES2,构建带有Twosc3基因的重组表达载体,转入酵母表达宿主菌,可生产得到木栓酮。通过突变研究结果表明,Twosc3基因编码的氨基酸482为关键位点,可以提高或降低木栓酮或香树素的产量。通过基因枪介导的干扰实验表明,Twosc3基因的干扰对于雷公藤中雷公藤红素的合成有明显的抑制作用,本发明TwOSC3蛋白及其编码基因可用于生物合成植物三萜类化合物,以及培育高品质的雷公藤。
权利要求
1.分离的蛋白,所述蛋白为
(1)SEQ ID NO:2所示的氨基酸序列;或
(2)SEQ ID NO:2所示的氨基酸序列经取代、缺失或增加一个或多个氨基酸且功能相同的肽。
2.根据权利要求1所述的蛋白,其中所述蛋白为将SEQ ID NO:2所示氨基酸序列中的一个或多个氨基酸进行突变:(1)将第482位的Leu突变为Ile、Ser、Val、Phe、Pro、Arg或Ala。
3.编码权利要求1或2所述蛋白的多核苷酸。
4.根据权利要求3所述多核苷酸,所述多核苷酸为如下中的至少一种:
(1)SEQ ID NO:1第115-2406位所示的核苷酸分子;或
(2)SEQ ID NO:1第115-2406位所示的核苷酸分子经取代、缺失或增加一个或多个核苷酸且表达相同功能蛋白的核苷酸序列;或
(3)在严谨条件下与SEQ ID NO:1第115-2406位所示核苷酸分子杂交的核苷酸序列,所述严谨条件为:在含0.1%SDS的0.1×SSPE或含0.1%的SDS的0.1×SSC溶液中杂交。
5.表达载体,其包含启动子、权利要求3或4所示多核苷酸和转录终止子,其中所述启动子与所述多核苷酸可操作地连接,并且所多核苷酸与所述转录终止子可操作地连接。
6.重组宿主细胞,其包含权利要求3或4所述多核苷酸分子、或权利要求5所述表达载体,所述细胞选自:细菌、酵母细胞、真菌细胞、昆虫细胞、哺乳动物细胞和植物细胞。
7.权利要求6所述细胞,其中所述植物细胞为雷公藤细胞。
8.权利要求1或2所述蛋白、或权利要求3或4所述多核苷酸分子在调节和生产植物三萜类化合物中的运用。
9.权利要求8所述的运用,其中所述三萜类化合物为木栓酮、β-香树素或雷公藤红素。
10.权利要求1或2所述蛋白、或权利要求3或4所述多核苷酸分子在雷公藤植物育种中的运用。
说明书
技术领域
本发明涉及一种雷公藤三萜合酶及其编码基因,以及该三萜合酶及其编码基因在三萜类化合物生物合成中的运用,属于药用植物基因工程领域。
背景技术
中药雷公藤来源于药用植物雷公藤(Tripterygium wilfordii.Hook.f.)的干燥根的木质部,被广泛用于类风湿性关节炎和炎症的治疗(Raphaela G M,Mildred W,Roy F,et al.Comparison of Tripterygium wilfordii Hook F Versus Sulfasalazine in theTreatment of Rheumatoid Arthritis:A Randomized Trial[J].Annals of InternalMedicine,2009,151(4):229-240.Tao X L,Lipsky P E.The Chinese anti-inflammatoryand immunosuppressive herbal remedy Tripterygium wilfordii Hook F.[J].Rheumatic Disease Clinics of North America,2000,26(1):29–50.)。萜类成分为雷公藤的主要活性成分,包括雷公藤甲素(triptolide)、雷酚内酯(triptophenolide)和雷公藤红素(celastrol)等。从中药中的活性成分开发新药是一种很有潜力的方式,然而由于植物的生长缓慢,再加上这些有效成分在植物体中的含量不多,因而大大限制了它的发展。通过探寻和阐释萜类成分在雷公藤中的生物合成途径及其调控机制,有助于为药材品质的形成提供理论基础,同时为利用生物技术提高目标成分含量或直接生产有效成分或中间体带来广阔的应用空间。
雷公藤中以木栓烷型的雷公藤红素为代表的三萜类化合物具有抗炎、抗风湿、抗肿瘤、减肥、免疫抑制、治疗神经性疾病等活性(Liu J,Lee J,Salazarhernandez M A,etal.Treatment of Obesity with Celastrol[J].Cell,2015,161(5):999–1011.)。通过胞浆的甲羟戊酸途径(mevalonic acid(MVA)pathway)和质体的2-methyl-D-erythritol-4-phosphate(MEP)途径生成萜类的通用底物异戊烯焦磷酸(Isopentenyl pyrophosphate(IPP))及其异构体Dimethylallyl pyrophosphate(DMAPP)。再由此分别生成单萜(monoterpenes),倍半萜(sesquiterpenes),二萜(diterpenes)和三萜(triterpenes)的底物香叶基焦磷酸(Geranyl diphosphate(GPP)),法尼基焦磷酸(farnesyl diphosphate(FPP))和牻牛儿基牻牛儿基焦磷酸(geranylgeranyl diphosphate(GGPP))。
三萜合酶(Triterpene synthase),又称三萜环化酶(Triterpene cyclase)或2,3-氧化鲨烯环化酶(2,3-oxidosqualene cyclase,OSC),被认为是合成三萜类次生代谢终产物的关键酶,它能够催化2,3-氧化鲨烯形成三萜骨架。本发明从雷公藤悬浮细胞中克隆得到一条Twosc3基因,通过验证其具有催化2.3-氧化鲨烯生成木栓酮的生物学功能,而木栓酮为木栓烷型三萜(雷公藤红素、扁塑藤素等)的母核结构,木栓酮经过修饰酶如P450等基因的修饰可以生成雷公藤红素等具有显著药理活性的三萜类化合物。该基因是首次从雷公藤中得到的三萜类成分合成的关键酶基因,在本发明被公布之前,尚未有任何公开或报道过本专利申请中所提及的雷公藤三萜合酶基因及其氨基酸序列。
发明内容
本发明提供了一种分离的蛋白,所述蛋白参与三萜类化合物的生物合成,尤其是木栓烷型三萜类化合物的生物合成。
本发明中,所述分离的蛋白为一种参与雷公藤三萜类化合物的合成,尤其是雷公藤红素合成的关键酶,本文将其命名为雷公藤三萜合酶(Tripterygium wilfordiiTriterpene synthase,TwOSC3),所述雷公藤三萜合酶具有:
(1)SEQ ID NO:2所示的氨基酸序列;或
(2)SEQ ID NO:2所示的氨基酸序列经取代、缺失或增加一个或多个氨基酸且功能相同的肽。
本发明中,TwOSC3蛋白或多肽,是指:具有三萜合酶活性(参与木栓烷型三萜类化合物的生物合成)的SEQ ID NO:2序列的多肽,当然还包括具有与天然TwOSC3相同功能的SEQ ID NO:2序列的变异形式。这些变异形式包括(但不限于):若干个(通常为1-50个,较佳地1-30个,更佳地1-20个,最佳地1-10个)氨基酸的缺失、插入和/或取代,以及在C末端和/或N末端添加一个或数个(通常为20个以内,较佳地为10个以内,更佳地为5个以内)氨基酸。例如,在本领域中,用性能相近或相似的氨基酸进行取代时,通常不会改变蛋白的功能。又比如,在C末端和/或N末端添加一个或数个氨基酸通常也不会改变蛋白质的功能。TwOSC3蛋白或多肽还包括TwOSC3的活性片段和活性衍生物,还包括能够可操作地连接于信号肽、启动子、核糖体结合位点或终止子序列组成的衍生物。
TwOSC3蛋白变体或具有实质上相似的序列一致性的多肽的特征在于具有一个或多个氨基酸替换、缺失或插入。这些改变优选在性质上较小,即保守氨基酸替代(见表1)和不显著影响多肽的折叠或活性的其它替代;小的缺失,典型地一个至约30个氨基酸的缺失;氨基或羧基端延伸,例如氨基端甲硫氨酸残基,不超过约20-25个残基的小的接头肽或亲和标签。本发明因此提供包含与SEQ ID NO:2至少有70%,优选至少有90%、更优选95%、96%、97%、98%、99%或更多一致的序列的多肽。
表1保守氨基酸替代
可以确定包含对于维持结构完整性关键的区域或域的氨基酸残基。可以确定在这些区域中或多或少耐受改变而保持分子的整个三级结构的特定残基。分析序列结构的方法包括,但不限于具有高氨基酸或核苷酸一致性的多个序列的比对、二级结构倾向(propensity)、二级元图(binary patterns)、互补堆积(complementary packing)和隐蔽的极性相互作用(Barton,Current Opin.Struct.Biol.5:372-376,1995;和Cordes等,Current Opin.Struct.Biol.6:3-10,1996)。一般地,当设计分子修饰或鉴定特定片段时,将在确定结构的同时评价修饰的分子的活性。
可以在TwOSC3蛋白中进行氨基酸序列改变以便最少破坏对生物学活性必须的高级结构。例如,当TwOSC3蛋白包含一个或多个螺旋时,将改变氨基酸残基以便不破坏螺旋的几何学和其中的构象改变将使一些关键功能(例如分子与其结合伙伴的结合)减弱的其它分子成分。氨基酸序列改变的效果可以通过例如以上公开的计算机模拟进行预测,或通过晶体结构分析进行测定(见例如Lapthorn等,Nat.Struct.Biol.2:266-268,1995)。本领域熟知的其它技术对改变蛋白和标准分子(例如天然蛋白)的折叠进行比较,例如,可以比较变体和标准分子中的半胱氨酸分布情况。质谱和使用还原和烷基化的化学修饰为测定与二硫键相关的或未形成这些键的半胱氨酸残基提供了方法(Bean等,Anal.Biochem.201:216-266,1992;Gray,Protein Sci.2:1732-1748,1993;和Patterson等,Anal.Chem.66:3727-3732,1994)。一般认为如果修饰的分子与标准分子具有不相同的半胱氨酸分布情况,则折叠受到影响。另一个用于测量这点的被接受的熟知方法是园二色谱(CD)。测量和比较修饰分子和标准分子产生的CD谱是常规的(Johnson,Proteins 7:205-214,1990)。晶体学是另一个分析折叠和结构的熟知方法,核磁共振(NMR)、消化肽作图和表位作图也是分析折叠及蛋白质和多肽之间的结构相似性的已知方法(Schaanan等,Science 257:961-964,1992)。
本发明中,雷公藤三萜合酶蛋白的变异体形式包括:同源序列、保守性变异体、等位变异体、天然突变体、诱导突变体、在高或低的严谨条件下能与雷公藤三萜合酶基因杂交的DNA所编码的蛋白。
优选的是,所述分离的蛋白变体为将SEQ ID NO:2所示氨基酸序列中的一个或多个氨基酸进行如下突变:
将第482位的Leu突变为Ile、Ser、Val、Phe、Pro、Arg或Ala。
本发明再一目的提供了一种编码所述雷公藤三萜合酶蛋白的多核苷酸序列。
所述的多核苷酸序列,是一种与雷公藤三萜类化合物合成有关的基因:即雷公藤三萜合酶基因(Twosc3基因),它是下列核苷酸序列之一:
1)序列表中SEQ ID No.1第115-2406位的核苷酸序列;或
2)与序列表中SEQ ID No.1第115-2406位限定的核苷酸序列具有一个或几个碱基突变,且编码相同功能蛋白质的DNA序列;或
3)在严谨条件下与SEQ ID NO:1第115-2406位所示核苷酸序列杂交的核苷酸序列,所述严格条件为:在含0.1%SDS的0.1×SSPE或含0.1%的SDS的0.1×SSC溶液中杂交,并用该溶液洗膜。
其中2404-2406的核苷酸序列“TAG”为终止密码子。
本发明所述雷公藤三萜合酶编码基因是指:编码具有雷公藤三萜合酶蛋白的核苷酸序列,如SEQ ID NO:1中第115-2406位核苷酸序列及其简并序列。该简并序列指位于SEQID NO:1序列的编码框第115-2406位核苷酸中,有一个或多个密码子被编码相同氨基酸的简并密码子所取代后而产生的序列。由于密码子的简并性,所以与SEQ ID NO:1中第115-2406位核苷酸序列同源性低至约70%的简并序列也能编码出SEQ ID NO:2所述的序列。还包括能在中度严谨条件下,更佳的在高度严谨条件下与SEQ ID NO:1中从核苷酸115-2406位的核苷酸序列杂交的核苷酸序列。还包括与SEQ ID NO:1中从核苷酸第115-2406位的核苷酸序列的同源性至少70%,较佳地至少80%,更佳地至少90%,最佳地至少95%的核苷酸序列。还包括能编码具有与天然的雷公藤三萜合酶相同功能的蛋白的SEQ ID NO:1中开放阅读框序列的变异形式。这些变异形式包括(但不限于)若干个(通常为1-90个,较佳地1-60个,更佳地1-20个,最佳地1-10个)核苷酸的缺失、插入和/或取代,以及在5’和/或3’端添加数个(通常为60个以内,较佳地为30个以内,更佳地为10个以内,最佳地为5个以内)核苷酸。
杂交反应的“严谨度”可以容易的由本领域普通技术人员确定,而且通常根据探针长度、洗涤温度、和盐浓度凭经验计算。通常,较长的探针要求较高的温度以正确退火,而较短的探针需要较低的温度。杂交通常依赖于当互补链存在于低于其解链温度的环境中时变性DNA重新退火的能力。探针和可杂交序列之间的期望同源性程度越高,可使用的相对温度也越高。结果是,推断出较高相对温度将趋向于使反应条件更为严格,而较低温度也就较不严格。关于杂交反应严谨度的另外细节和解释,参见Ausubel等人,《Current Protocols inMolecular Biology》,Wiley Interscience Publishers,1995。
“严谨条件”或“高严谨条件”,如本文中所定义的,可如下鉴别:(1)采用低离子强度和高温进行洗涤,例如0.015M氯化钠/0.0015M柠檬酸钠/0.1%十二烷基硫酸钠,50℃;(2)在杂交过程中采用变性剂,诸如甲酰胺,例如50%(v/v)甲酰胺及0.1%牛血清清蛋白/0.1%Ficoll/0.1%聚乙烯吡咯烷酮/50mM磷酸钠缓冲液pH 6.5,含750mM氯化钠,75mM柠檬酸钠,42℃;或(3)在采用50%甲酰胺,5x SSC(0.75M NaCl,0.075M柠檬酸钠),50mM磷酸钠(pH 6.8),0.1%焦磷酸钠,5x Denhardt氏溶液,超声波处理的鲑鱼精DNA(50μg/ml),0.1%SDS,和10%硫酸右旋糖苷的溶液中于42℃杂交过夜,以及于42℃在0.2x SSC(氯化钠/柠檬酸钠)中洗涤10分钟,接着在含EDTA的0.1x SSC中于55℃进行10分钟高严格性洗涤。
本发明的还一目的在于提供一种表达载体,所述表达载体包含编码本发明所述雷公藤三萜合酶蛋白或其变体的多核苷酸,或在严谨条件下可与所述多核苷酸杂交的核苷酸序列,所述表达载体还包含启动子和终止子,其中所述启动子与所述多核苷酸可操作地连接,并且所述多核苷酸与所述转录终止子可操作地连接。
本发明中,可选用本领域已知的各种载体,如市售的载体,包括质粒、粘粒等。在生产本发明雷公藤三萜合酶多肽时,可以将雷公藤三萜合酶基因的核苷酸序列可操作地连接于表达调控序列,从而形成雷公藤三萜合酶表达载体。所述“可操作地连连接”当指DNA区段时表示这些区段按照一定方式排列以致它们可以协调地为其预期的目的发挥作用,例如在启动子中起始转录并前行通过编码区段到达终止子。也指这样一种状况:即线性DNA序列的某些部分能够影响同一线性DNA序列其它部分活性,例如,如果信号肽DNA作为前提表达并参与多肽的分泌,那么信号肽(分泌前导序列)DNA就是可操作地连接接于多肽DNA;如果启动子控制序列转录,那么它是可操作地连接于编码序列;如果核糖体结合位点被置于能使其翻译的位置时,那么它是可操作地连于编码序列。一般,“可操作地连接”意味着相邻,而对于分泌前导序列则意味着在阅读框中相邻。
本发明的再一目的,在于提供一种宿主细胞,所述宿主细胞,包含本发明所述编码TwOSC3蛋白或其变体的多核苷酸分子,或在严谨条件下可与所述多核苷酸分子进行杂交的核苷酸分子,或包含本发明上述描述的表达载体。所述的宿主细胞选自:细菌、原核细胞(如大肠杆菌、)真菌细胞、酵母细胞、昆虫细胞、哺乳动物细胞或植物细胞,优选地,为酵母细胞或植物细胞。
特别有意义的酵母包括酿酒酵母、巴斯德毕赤酵母和Pichia methanolica。用外源DNA转化酿酒酵母细胞和从其中制备重组多肽的方法公开在例如Kawassaki,美国专利US4599311,US4931373、US4870008、US5037743、US4845075等中。通过选择标记所确定的表型,通常是药物抗性或在缺少特定养分(例如亮氨酸)时的生长能力,选择转化细胞。用于酿酒酵母的优选载体系统如可以是pYES2表达载体。用于酵母的适宜启动子和终止子包括来自糖酵解基因(US4599311、US4615974和US4977092)和醇脱氢酶的那些。用于其它酵母,包括多形汉逊氏酵母、乳克鲁维氏酵母、脆壁克鲁维氏酵母、巴斯德毕赤酵母、PichiaMethanolica、季也蒙氏毕赤酵母和麦芽糖假丝酵母的转化系统也是本领域已知的。
根据常规方法在含有养分和其它对于所选宿主细胞的生长必须的成分的培养基中培养转化的或转染的宿主细胞。多种适宜的培养基,包括已知成分培养基和复杂培养基,是本领域已知的,一般包括碳源、氮源、必需氨基酸、维生素和矿物质。如果需要,培养基还可以含有诸如生长因子或血清这样的成分。生长培养基一般通过例如药物筛选或缺少可由表达载体携带的或共转染至宿主细胞中的选择标记补充的必需养分来选择含有外源添加DNA的细胞。通过常规方式,如振摇小三角瓶或发酵罐喷气给液体培养物提供充足的空气。
本发明的雷公藤三萜合酶多核苷酸全长序列或其片段通常可以用PCR扩增法、重组法或人工合成的方法获得。对于PCR扩增法,可根据本发明所公开的有关核苷酸序列,尤其是开放阅读框序列来设计引物,并用市售的cDNA库或按本领域技术人员已知的常规方法制备cDNA库作为模板,扩增而得有关序列。当序列较长时,常常需要进行两次或多次PCR扩增,然后再将各次扩增出的片段按正确次序拼接在一起。一旦获得了有关的序列,就可以用重组法来大批量地获得有关序列。这通常是将其克隆入载体,再转入细胞,然后通过常规方法从增殖后的宿主细胞中分离得到有关序列。此外,还可通过化学合成将突变体引入本发明蛋白序列中。除了用重组法产生之外,本发明蛋白的片段还可用固相技术,通过直接合成肽而加以生产(Stewart等,Solid-Phase Pedtide Synthesis,J.Am.Chem.Soc.85:2149-2154,1963)。在体外合成蛋白质可以用手工或自动进行。例如,可以用Applied Biosystems的431A型肽合成仪(Foster City,CA)来自动合成肽。可以分别化学合成本发明蛋白的各片段,然后用化学方法加以连接以产生全长的分子。
本发明中,术语“雷公藤三萜合酶基因”或“Twosc3基因”可互换使用;术语“雷公藤三萜合酶”、“三萜合酶”、“雷公藤三萜合酶蛋白”、“TwOSC3”或“TwOSC3蛋白”可互换使用。术语“核苷酸分子”或“核苷酸序列”可互换使用。
术语“分离的”或“纯化的”,多肽或蛋白质是指在非其天然环境的条件下,例如离开血液或组织存在的多肽或蛋白质。在优选形式中,分离的多肽或蛋白基本上不含有其他多肽或蛋白,尤其是动物来源或植物来源的其它多肽或蛋白。优选提供高纯度化形式,即纯度大于95%、更优选纯度大于99%的多肽;“分离的”或“纯化的”DNA是指,该DNA或片段已从天然状态下位于其两侧的序列中分离出来,或指该DNA或片段已经与天然状态下伴随核酸的组分分开,而且已经在与细胞中伴随其的蛋白质分开。
本发明提供的雷公藤三萜合酶蛋白及雷公藤三萜合酶基因是首次从雷公藤中克隆制备的,其直接参与三萜类化合物的生物合成,典型地化合物如木栓酮。由于木栓酮为木栓烷型三萜类化合物(雷公藤红素)的前体结构,因此可以通过基因工程技术来提高植物中木栓酮的含量,或者木栓烷型三萜类化合物如雷公藤植物中雷公藤红素的含量,基因枪介导的Twosc3基因干扰结果显示,雷公藤三萜合酶基因的干扰对于雷公藤中雷公藤红素的合成有明显的抑制作用,转入含Twosc3基因的干扰载体的悬浮细胞对比转入空载的悬浮细胞Twosc3基因表达量显著下降(干扰),相对应的雷公藤红素含量显著下降(干扰),提示其对雷公藤红素合成具有重要作用。Twosc3基因可用于利用转基因技术来提高雷公藤红素含量的研究和产业化中,尤其可用于中药雷公藤的品质改良,对于缓解雷公藤药源匮乏问题有较好的促进作用,可用于雷公藤育种。
本发明的一个实施方案中,提供了本发明所述雷公藤三萜合酶或雷公藤三萜合酶基因在合成三萜类化合物中的运用,所述三萜类化合物,如可以是木栓酮、雷公藤红素等。
本发明的一个实施方案中,提供了一种通过运用本发明所述雷公藤三萜合酶或雷公藤三萜合酶基因调节和生产植物三萜类化合物的方法,所述三萜类化合物如木栓酮或雷公藤红素。运用TwOSC3蛋白或Twosc3基因获得植物三萜类成分的方法包括:(1)雷公藤悬浮细胞的培养;(2)雷公藤悬浮细胞RNA提取;(3)雷公藤悬浮细胞转录组测序;(4)Twosc3基因全长cDNA获取;(5)质粒和菌株构建;(6)工程菌培养;(7)工程菌表达产物提取和分离。
附图说明
图1 Twosc3基因的组织表达(每个样本三个生物学重复,三个技术重复)
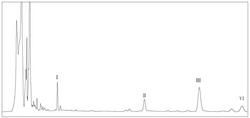
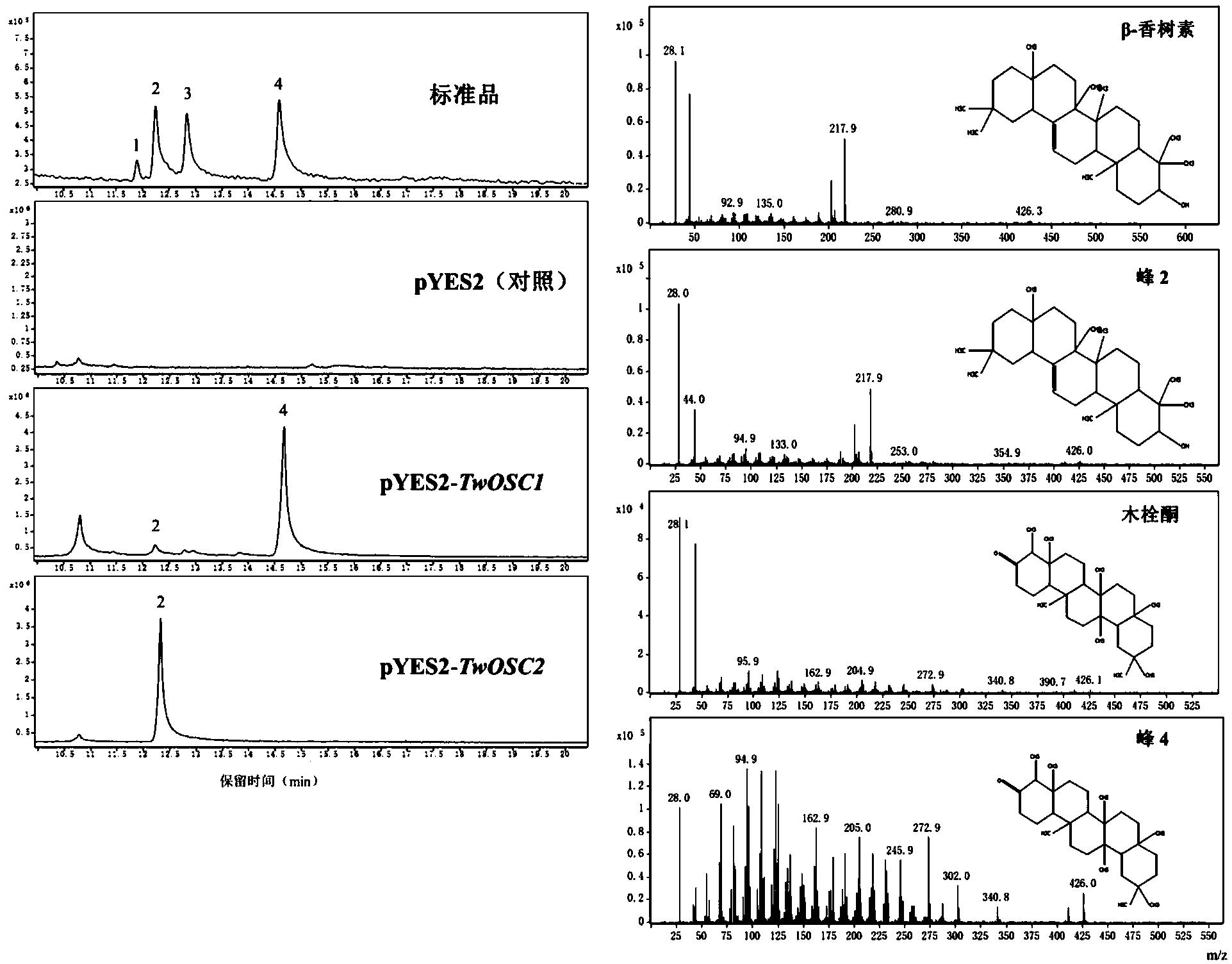
图2 Twosc3基因发酵产物的GC-MS结果图(标准品峰1为β-谷甾醇,2为β-香树素,3为羽扇豆醇,4为木栓酮)
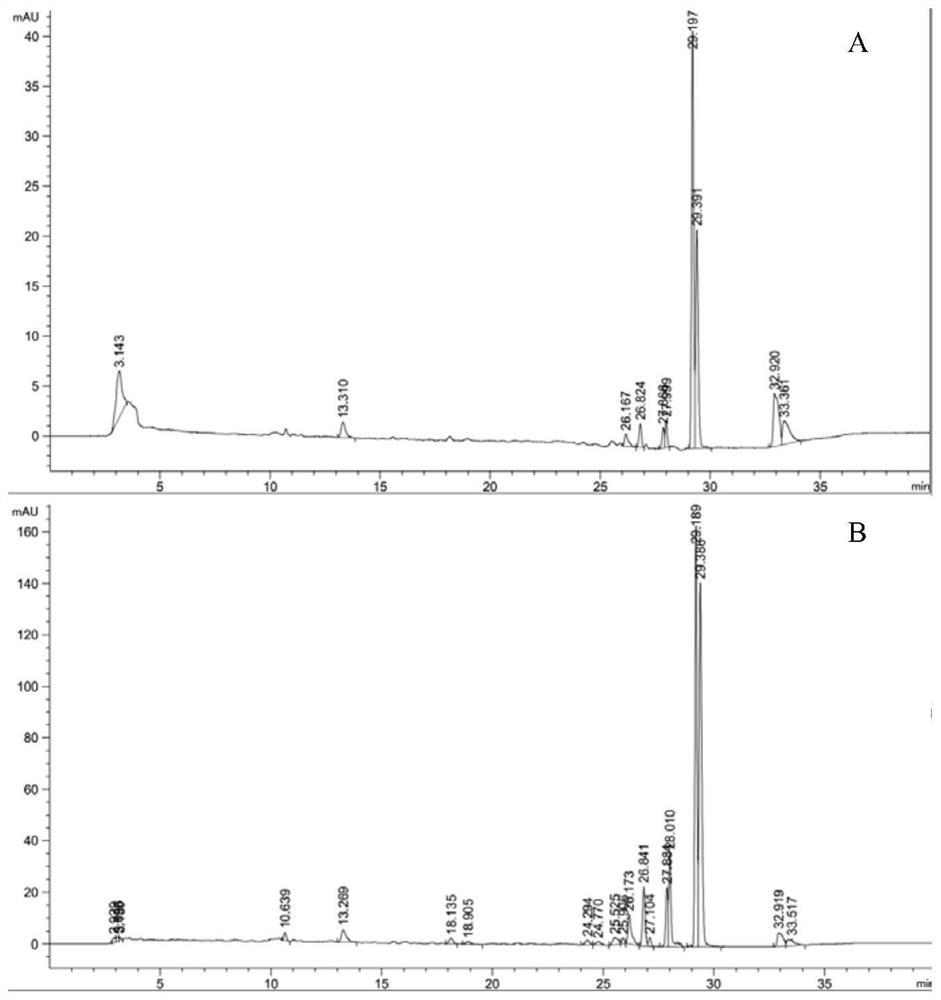
图3 TwOSC3蛋白482位突变结果产物GC-MS检测图(1为β-香树素,2为木栓酮)
图4 Twosc3及其突变体的产物含量测定结果
图5雷公藤悬浮细胞Twosc3基因干扰及雷公藤红素的含量(CK为转入空载的悬浮细胞,Ri为转入带Twosc3基因干扰载体的雷公藤悬浮细胞)
具体实施方式
下述实施例中所使用的实验方法如无特殊说明,均为常规方法。
下述实施例中所用的材料、试剂等,如无特殊说明,均可从商业途径得到。
下述实施例中的定量试验,均设置三次重复实验,结果取平均值。
下述实施例中的雷公藤(Tripterygium wilfordii Hook.f.)悬浮细胞在文献“雷公藤4-(5’-二磷酸胞苷)-2-C-甲基-D-赤藓醇激酶基因的全长克隆与表达分析.中国中药杂志,2015,40(21):4165-4170”中公开过,公众可从首都医科大学分子生药与中药资源实验室获得。
下述实施例中的pEASY-Blunt Simple Cloning Kit和Fast Mutagenesis System是北京全式金生物技术有限公司的产品;
Phusion High-Fidelity PCR Master Mix with HF Buffer和各种核酸内切酶均购自New England Biolabs公司;
SMARTerTM RACE cDNA Amplification Kit是Takara公司的产品;
FastQuant RT Kit试剂盒、快速质粒小提试剂盒均购自天根生化科技有限公司。
β-香树素(β-Amyrin)是美仑生物公司产品,产品批号为M0505AS,CAS号为559-70-6;
木栓酮(Friedelin)是美仑生物公司的产品,产品批号为M0308AS,CAS号为559-74-0;
羽扇豆醇(Lupeol)是阿拉丁(aladding)公司产品,产品批号为#JI409077,货号L114079-10mg,CAS号为545-47-1;
β-谷甾醇(β-sitosterol)是阿拉丁(aladding)公司产品,产品批号为#D1605025,货号S111183-20mg,CAS号为83-46-5。
实施例1、雷公藤Twosc3全长cDNA序列的克隆
1.雷公藤悬浮细胞总RNA提取及cDNA第一链的获得
采用改良CTAB法(CTAB Buffer:2%CTAB(W/V);100mmol·L
2.引物设计
根据雷公藤转录组数据注释筛选得到基因全长序列片段,设计Twosc3-F和Twosc3-R引物,引物序列如下:
Twosc3-F:AAACACAAGAGATTGATATAAGC(SEQ ID NO:3)
Twosc3-R:TTCGATTTAGCGGACACT(SEQ ID NO:4)
3.PCR扩增
以步骤1获得的5’-RACE-Ready cDNA为模板,采用Twosc3-F和Twosc3-R引物进行PCR扩增,得到PCR扩增产物。
PCR反应程序:98℃预变性30s;98℃10s,60℃15s,72℃2min,35个循环;72℃延伸5min。
扩增产物经Gene JET Gel Extraction Kit胶回收(方法如下)。
Gene JET Gel Extraction Kit胶回收步骤:
(1)取PCR产物与9μL的6×loading buffer预混合,在1.5%琼脂糖凝胶上以低电压(约5Vcm-1)电泳30-60min;
(2)用解剖刀或剃刀片切割含有DNA片段的凝胶,将胶片放在事先称重的1.5mL离心管并称重。记录胶片的重量。
(3)加1:1量的Binding Buffer到胶片中(量以重量计,如每100毫克琼脂糖凝胶加100微升的Binding Buffer);
(4)在50-60℃的条件下温育凝胶混合物10min,期间颠倒混匀2-3次,促进胶融化,保证胶全部溶解,在上柱前将凝胶混合物快速涡旋混匀一次;
(5)转移最多800μL凝胶溶解液到基因回收纯化柱,13000g离心1min,弃去流出液,然后将柱放回相同的收集管;
(6)加入700μLWash Buffer(已用乙醇稀释)到Gene JET纯化柱。13000g离心1min,弃去流出液,然后将柱放回相同的收集管;
(7)离心空GeneJET纯化柱13000g离心1min,彻底去除残留的Wash Buffer;
(8)将Gene JET纯化柱转移到一个干净的1.5mL离心管,加30-50μL ddH2O(可60℃预热)于纯化柱膜,13000g离心1min;
(9)丢掉Gene JET纯化柱并储存纯化的DNA在-20℃。
3.载体连接
取4μL的回收产物,加入1μLpEASY-Blunt Simple Cloning Kit试剂中的B-Zero预混液,混匀后,在PCR仪中25℃,连接15min。
4.连接产物转化
(1)将上述连接产物全部加入到50μL的Trans1-T1感受态细胞中,冰中放置30min;
(2)42℃热激1min后,再放置冰中2min;
(3)加入1mLLB培养基,37℃,180rpm振荡培养1h;
(4)吸取200μL菌液涂在LB+Amp固体培养基上,37℃培养14h,观察菌落生长情况。
5、菌液PCR验证阳性菌落
在超净台中,用灭过菌的10μL白枪头挑取平板中的单一菌落,置于LB+Amp培养基中,37℃,250rpm恒温振荡培养约1-2h后,取1μL菌液进行菌液PCR,检测目的片段是否与载体有效连接上。
PCR反应体系:
PCR反应条件:
PCR产物经过1%琼脂糖凝胶电泳检测后,凝胶成像仪上观察。将出现2200bp左右目的片段条带的菌液送测序公司测序;
测序结果表明:PCR扩增产物的序列如序列1所示,将序列1所示的基因命名为Twosc3,其中,自5’端第115-2406位为开放阅读框(ORF),编码由763个氨基酸残基组成的蛋白质(其中2404-2406的核苷酸序列“TAG”为终止密码子),该蛋白命名为TwOSC3,该蛋白的氨基酸序列为序列2。
实施例2、Twosc3基因组织表达分析
1.实验材料的处理
雷公藤根、茎、叶、花采自于福建永安国有林场五个不同植株。样品取回实验室清洗,液氮速冻后保存于负80冰箱。
2.总RNA的提取及Real-time定量PCR
将存于负80冰箱的根、茎、叶、花在液氮环境下粉碎,用改良CTAB法提取总RNA,用FastQuant RT Kit试剂盒(天根)反转录成cDNA,以雷公藤EF1α基因作为内参。采用ABIPrism 7300Sequence Detection System(Applied Biosystems,USA)和KAPA SYBR FASTUniversal 2X qPCR Master Mix(Kapa Biosystems,USA)试剂盒进行Real-time定量PCR。实时荧光定量反应体系:2×Fast qPCR Master Mix 10uL,正反引物(10mmol·L-1)各0.4uL,cDNA1uL,50×Rox High 0.4uL,PCR-grade water 7.8uL。反应条件为:95℃3min,95℃3s,60℃30s,45个循环:65~95℃做溶解曲线分析。反应结束后对荧光值扩增曲线和融解曲线进行分析。生物重复3个,技术重复3次,采用2
Twosc3-F:5’-AGTATCCTGATGGCTCGTGG-3’(SEQ ID NO:5)
Twosc3-R:5’-CCCATCCACCATCATCACTC-3’(SEQ ID NO:6)
EF1α-F:5’-CCAAGGGTGAAAGCAAGGAGAGC-3’(SEQ ID NO:7)
EF1α-R:5’-CACTGGTGGTTTTGAGGCTGGTATCT-3’(SEQ ID NO:8)
结果如图1所示。经EF1α内参基因均一化后,Twosc3基因相对表达水平最高的是在根中,其次是茎,然后是花,Twosc3基因相对表达量最低在叶中。提示Twosc3主要负责根中木栓酮的合成,这与雷公藤红素主要分布部位一致。
实施例3、雷公藤Twosc3生物学功能研究
1.真核表达载体构建
以含有雷公藤Twosc3基因全长cDNA的载体pEASY-Blunt-Twosc3质粒为模板,用含酶切位点引物(标横线的为酶切位点),进行PCR扩增基因编码区。DNA聚合酶采用高保真DNA聚合酶(Phusion High-Fidelity PCR Master Mix)。PCR参数为98℃30s,1循环;98℃10s,60℃10s,72℃2min 30s,35循环;72℃5min;4℃维持。扩增产物经Gene JET GelExtraction Kit胶回收(方法如下)。
Twosc3-F:CGG
Twosc3-R:CGC
Gene JET Gel Extraction Kit胶回收步骤(同实施实例1中胶回收步骤):
经回收后的PCR产物用限制性内切酶进行双酶切,采用NEB公司T4 DNA连接酶定向连入经相同双酶切的表达载体pYES2中(方法如下)。
双酶切反应体系(50μL体系)
37℃反应2h后琼脂糖凝胶电泳,Gene JET Gel Extraction Kit胶回收目的片段方法同上。
连接反应(20μL体系)
*DNA与载体摩尔比约为3:1—10:1
25℃连接1h,连接产物经转化、阳性克隆的初步筛选(具体步骤同实施实例1中连接产物的转化和单菌落菌液PCR验证),送样测序鉴定,得到经测序核苷酸序列无突变重组质粒pYES2-Twosc3。
2.酵母发酵
酵母感受态的制备
将ATCC处购买的羊毛甾醇合酶缺陷酵母( 4021900
(1)菌液在低温离心机500g,离心4min,弃上清。
(2)加入10mL EZ 1solution洗涤菌体,500g离心4min,弃上清。
(3)加入1mL EZ 2solution重悬菌体,每50uL菌液分装到1.5mL EP管中,梯度降温后保存在负80℃冰箱或直接用于载体转化。
(4)取0.2-1ug的质粒(少于5uL)加入50uL酵母感受态细胞,加500uL EZ3solution,完全混匀。
(5)30℃孵育45min。期间用手指轻弹或斡旋2-3次。
(6)取50uL-150uL孵育液涂布在Sc-ura酵母筛选培养基上,30℃暗培养2d。
发酵流程
空载转化的酵母菌作为对照,挑取单菌落于5mL Sc-ura液体培养基30℃,250rpm培养过夜,用于菌液PCR验证。将含有目的条带的菌保存为甘油菌,并取20uL菌液在20mL新的Sc-ura液体培养基30℃,250rpm培养2d,换为Sc-ura诱导培养基(含半乳糖),30℃,220rpm诱导培养12h,再用20mL的0.1M磷酸钾(含葡萄糖,PH7.0)30℃,220rpm培养1d。
3.产物GC-MS检测
发酵后的菌液离心,取菌体加入10mL 20%KOH/50%EtOH溶液超声提取10min,用10mL正己烷萃取三次,萃取液合并后,氮吹挥去溶剂,加入100uL无水吡啶和100uL三氟乙酰胺(含三甲基硅烷)65℃衍生化60min。氮吹挥干溶剂后,800uL氯仿复溶,过滤膜,GC-MS检测。GC-MS条件:进样量1uL,50℃保持1min,50℃·min
结果见图2。与转入空载的对照组酵母提取物相比,转入Twosc3基因的酵母提取物中检测到木栓酮,说明Twosc3基因表达的蛋白能够催化2,3-氧化鲨烯形成木栓酮。
4.定点突变
除突变位点外,两条引物长度大约25-30bp,5’端重叠区包含15-20bp,3’端延伸区包含至少10bp;突变位点位于两条引物上,分别位于正向突变引物重叠区下游、紧邻重叠区,反向突变引物5’端根据以上原则设计定点突变引物,在‘DCTAE’功能结构域上游第二个氨基酸处(482位)将Twosc3CTT(L)突变成GTT(V),ATT(I),TCT(S),TTT(F),GCT(A),CCT(P)和CGT(R)。定点突变引物序列:
反应体系
上述体系混匀后,与PCR仪中,预变性94℃5min,94℃20s,55℃20s,72℃3min 30s,25个循环,72℃延伸10min。反应结束后加入1uL DMT酶消化产物,37℃反应1h。取4uL消化产物加入50uL DMT感受态细胞,混匀,冰浴30min,42℃水浴45s,冰浴2min,加入500uL的LB培养基,200rpm,37℃培养1h,涂布于LB+AMP培养皿上37℃培养。阳性克隆送测序成功后,提取质粒。
将提取的突变质粒转入酵母感受态细胞中(实例3中步骤2制备),于Sc-ura酵母培养基培养,随后挑取单菌落进行发酵和诱导,提取产物,GC-MS检测结果(步骤同实施实例3中的2和3)。
结果如图3所示。在Twosc3编码的氨基酸序列上第482位的亮氨酸(L)分别突变成丙氨酸(A)、苯丙氨酸(F)、精氨酸(R)和脯氨酸(P)后产物均消失,说明酶的活性丧失;而482位分别突变成缬氨酸(V)、丝氨酸(S)后产物变为2(β-香树素),产物4(木栓酮)消失,其中将Twosc3编码的氨基酸序列上第482位的亮氨酸(L)突变成异亮氨酸(I)后,同时生成产物2和产物4,且产物4的含量有所提升。进一步将产物含量有变动的突变体菌株以及Twosc3野生型菌株分别进行发酵定量,发酵步骤及产物提取检测方法均同上。每个菌株三个重复,产物测定结果见图4。
实施例4、雷公藤Twosc3基因干扰对雷公藤红素合成的影响
1.雷公藤悬浮细胞准备
配制MS固体培养基:Murashige&Skoog with Vitamins(Caisson Labs公司)4.43g/L,蔗糖(AOBOX公司)30g/L,2,4-D 1.0mg/L,KT 0.1mg/L,加水后调PH到5.8-6.0,加琼脂糖(Oxoid公司)6-8g,121℃,20min高压灭菌后温度降到60℃以下时倒入35mm细胞培养皿(每个培养皿3ml培养基)中凝固。
将要继代的悬浮细胞转入加入培养基的35mm细胞培养皿中(每个培养皿0.2g悬浮细胞),25℃,暗培养6-9d用于基因枪转化。
2.gateway技术构建Twosc3干扰载体
BP入门载体构建
设计引物如下:
干扰引物Twosc3-F:
Twosc3-R:CATACCACGAGCCATCAGG(SEQ ID NO:26)
以含有雷公藤Twosc3基因全长cDNA的载体pEASY-Blunt-Twosc3质粒为模板,用含酶切位点引物(标横线的为酶切位点),进行PCR扩增基因编码区。DNA聚合酶采用高保真DNA聚合酶(Phusion High-Fidelity PCR Master Mix)。PCR参数为98℃30s,1循环;98℃10s,60℃10s,72℃2min 30s,35循环;72℃5min;4℃维持。扩增产物经Gene JET GelExtraction Kit胶回收。
BP反应体系
22℃反应2h后转入大肠杆菌感受态细胞Trans1-T1(北京全式金公司)于LB+Kan培养基37℃培养,阳性克隆送测序成功后,快速质粒小提试剂盒提取质粒。
LR反应体系
*pK7为干扰载体
25℃反应3-4h,加入1uL蛋白酶K,37℃温育10min,转化入Trans-T1于LB+Spe培养基37℃培养,阳性克隆送测序成功后,用Plasmid Maxi Kit(Omega公司)质粒大提试剂盒提取质粒(质粒浓度要求>1ug/uL)。
3.基因枪介导转化
实验器材要求无菌:根据器材性质分别或结合采用高温高压灭菌、紫外照射除菌和70%乙醇杀菌。
每个质粒做5个生物学重复,即打5盘细胞,每盘2枪,共10枪。将转化pK7空载体的悬浮细胞用作对照。将20ug质粒,5000ug金粉,100uL 2.5M的CaCl2,40uL 0.1M的亚精胺混合,震荡2-3min,静止1min后离心约2s,弃上清;加入140uL 70%乙醇,轻弹后离心弃上清;加入140uL100%乙醇,轻弹后离心弃上清;加入100uL100%乙醇,轻弹后,在低速震荡2-3s,置于冰中。
将消毒后的载体膜置于铁托中,每个铁托加入10uL上述配制的载体金粉预混液,按基因枪操作规范,将铁托,阻挡网,可裂膜及雷公藤悬浮细胞分别放置在基因枪设备的相应位置上,进行基因枪转化载体。每盘细胞打两枪。
4.筛选培养及基因表达分析
将基因枪转化完的细胞,25℃,暗培养2d,转入MS筛选培养基为MS+Kan培养基,抗生素终浓度Kan为100mg/ml。20d筛选完成后,将筛选后的细胞继代于新的MS+Kan的培养基中继续培养20d,之后转移到MS+Kan的液体培养基中,25℃,120rpm培养3代后,取一部分细胞液氮速冻后用于提取RNA,剩余细胞真空冷冻干燥后,取0.2g甲醇1ml超声2h提取产物,UPLC分析产物结果。
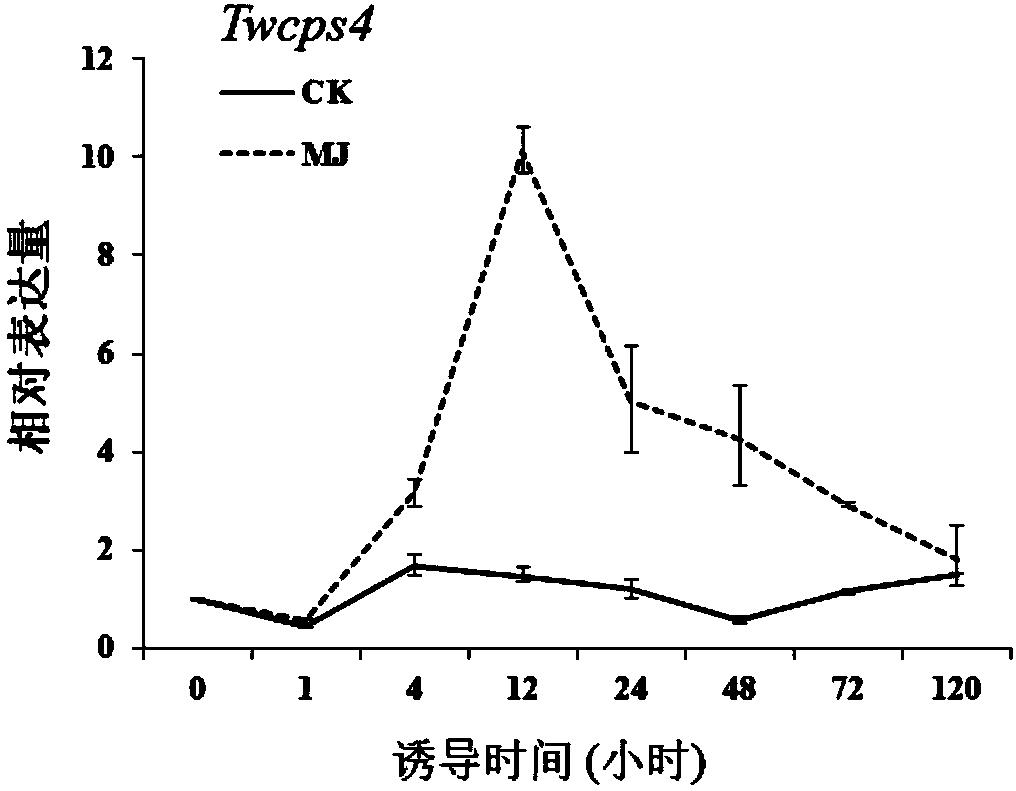
液氮速冻后的细胞,在液氮环境下粉碎,用RNA提取试剂盒(Promega公司)提取总RNA,用FastQuant RT Kit试剂盒(天根)反转录成cDNA,以雷公藤EF1α基因作为内参。采用ABI Prism 7300Sequence Detection System(Applied Biosystems,USA)和KAPA SYBRFAST Universal 2X qPCR Master Mix(Kapa Biosystems,USA)试剂盒进行Real-time定量PCR。实时荧光定量反应体系:2×Fast qPCR Master Mix 10uL,正反引物(10mmol·L-1)各0.4uL,cDNA1uL,50×Rox High 0.4uL,PCR-grade water 7.8uL。反应条件为:95℃3min,95℃3s,60℃30s,45个循环:65~95℃做溶解曲线分析。反应结束后对荧光值扩增曲线和融解曲线进行分析。生物重复5个,技术重复3次,采用2
结果见图5。转入含Twosc3片段的干扰载体的悬浮细胞对比转入空载的悬浮细胞Twosc3基因表达量显著下降,相对应的雷公藤红素含量显著降低(11.73ug/g比空载下降了77%)。结果显示Twosc3的干扰可以下调雷公藤红素的合成,表明与雷公藤红素的生成显著相关。
上述说明并非对本发明的限制,本发明也并不限于上述举例。本技术领域的普通技术人员在本发明的实质范围内,作出的变化、改型、添加或替换,也应属于本发明的保护范围,本发明的保护范围以权利要求书为准。
序列表
<110> 首都医科大学
<120> 雷公藤三萜合酶TwOSC3及其编码基因与应用
<130> 背景技术中
<141> 2018-08-08
<160> 26
<170> SIPOSequenceListing 1.0
<210> 1
<211> 2475
<212> DNA
<213> Abracris flavolineata
<400> 1
aaacacaaga gattgatata agctctcttg tcttagttcc tcatcatatc atcaagttct 60
tggaggcttc aacctctcat ccttgatagg cttcctctct ggagtgaaga aatcatgtgg 120
aagataaaga tagctgaacg tgggaactgc ccctactccg agtacttgta cactacaaat 180
gactttgttg gtaggcagat atgggagttt gaccctaatg caggcacccc tgaagagctc 240
gctgaggtcg aagaggctcg caagaaattc accgagaacc gccatgaagt caagcctgct 300
tccgacctac tttggatgca gcagtttctg aggcagaaga acttcaagca aacaattcct 360
ccgttgagaa ttggggagaa agaacaagtt acatatgaag atgtcacaac tgctttgaag 420
agagcttcct ctttcttgtc agccttgcaa gccagtgatg gtcactggcc agcagaaaat 480
gctggagtct tgtttttcct tcctccattt atcttttgcc tatacatcac tggacatctc 540
aattcgatta tcacttcgga acatcgtaaa gaaatcctac gtttcatata caaccaccag 600
aatgaagatg ggggttgggg aatacacata gagggacaca gctcaatgtt taccacaaca 660
ttcagctatg tgtgcatgag aatacttgga gtaggaccag atgaaagtgc ttgtgctaga 720
gcaagaaagt ggattcttga tcatggtggc atcacttaca tggcttcctg gggaaagacc 780
tggctttcga tacttggtat atttgattgg tatggatgca acccaatgcc cccagagttc 840
tggattcttc cttcttatct ccctatacat ccagcgaaaa tgtggtgcta ttgccggatg 900
gtttacatgc caatgtcata tctatatggg aaaagatttg ttggtcctat cacacctctc 960
attttacaat tgagagagga gctccacact cagccctacc atgaaattca atggagacaa 1020
atgcgccatc gatgcggaaa ggaggatcta tactatcccc acagtctaat ccagaatttc 1080
ctctgggacg gtctttatgt ggcttctgaa cctcttttga ctcgctggcc ttttagcaag 1140
ataagagaga aggctcttga aaaagcaatg gaacacattc actacgaaga cgaaaatagt 1200
cgatacatca ccattggctg tgttgaaaag gcattgtgta tgctttgttg ctgggttgaa 1260
gaccccaatg gagactattt caagaagcat cttgcaagga taccagatta cttatgggtt 1320
gcagaggatg gtatgaaagt ggtgagtttt ggcagccaac tgtgggatgc tacctttggc 1380
tttcaagctc tggttgctag caatctaaca aatgaagtcg ctccaactct cgtcaaagca 1440
tacgacttcg taaagaaatg ccaggtccgg aataatccct ctggtgactt tgagaaaatg 1500
tttcgccaca tttcaaaagg atcatggact ttctccgatc aagatcatgg atggcaactt 1560
tctgattgca ctgctgaagc attaaagtgt tgcttgctag cagcaaccat gcccgaagaa 1620
cttgttggag agaaattaga tcctcaatgg atatttgagt ctgtaaacat catactttcc 1680
ctccaagagc ctaaaaccgg cggtttagct ggctgggaac cagtaagagc tggacaatgg 1740
atggagatgc tcaaccctat ggagtttctc gagaatattg tgattgaaca tacatatgtc 1800
gagtgtactg gatcttccat cattgctttc attacactca agaagttatt cccaggtcat 1860
aggaccaaag acattgacaa tttcattgta aacggtataa gatatctcga ggacgtgcag 1920
tatcctgatg gctcgtggta tggaaactgg ggtatttgct tcatctatag taccatgttt 1980
gcacttggag ggctagcagc agctggtagg acttacgaaa attgtcccgc tgtgcgcaga 2040
ggtgttgatt ttctacttat taatcagagt gatgatggtg gatggggaga aagctatctt 2100
tcttccccaa gaaagatata tacacctctt gaaggaagaa gatcaaatgt ggtacaaact 2160
gcatgggcta tgctagggct gctctatgct ggccaggttg agagagatcc aactcctctt 2220
caccgcggtg cgaagatact aatcaattac caaatggaag agggaggtta cccccagcag 2280
gaaatcactg gagttttcaa aatgaattgc atgttacact atccaatata taggaatgcc 2340
ttcccaatat gggcgctcgg agagtatcga aatcgtgtta agttgccttc aaaggaaatt 2400
aaatagcaat gaaactaaat tccgtgtaaa aagacgtgaa tgagaataat gtacacaagt 2460
gtccgctaaa tcgaa 2475
<210> 2
<211> 763
<212> PRT
<213> Abracris flavolineata
<400> 2
Met Trp Lys Ile Lys Ile Ala Glu Arg Gly Asn Cys Pro Tyr Ser Glu
1 5 1015
Tyr Leu Tyr Thr Thr Asn Asp Phe Val Gly Arg Gln Ile Trp Glu Phe
202530
Asp Pro Asn Ala Gly Thr Pro Glu Glu Leu Ala Glu Val Glu Glu Ala
354045
Arg Lys Lys Phe Thr Glu Asn Arg His Glu Val Lys Pro Ala Ser Asp
505560
Leu Leu Trp Met Gln Gln Phe Leu Arg Gln Lys Asn Phe Lys Gln Thr
65707580
Ile Pro Pro Leu Arg Ile Gly Glu Lys Glu Gln Val Thr Tyr Glu Asp
859095
Val Thr Thr Ala Leu Lys Arg Ala Ser Ser Phe Leu Ser Ala Leu Gln
100 105 110
Ala Ser Asp Gly His Trp Pro Ala Glu Asn Ala Gly Val Leu Phe Phe
115 120 125
Leu Pro Pro Phe Ile Phe Cys Leu Tyr Ile Thr Gly His Leu Asn Ser
130 135 140
Ile Ile Thr Ser Glu His Arg Lys Glu Ile Leu Arg Phe Ile Tyr Asn
145 150 155 160
His Gln Asn Glu Asp Gly Gly Trp Gly Ile His Ile Glu Gly His Ser
165 170 175
Ser Met Phe Thr Thr Thr Phe Ser Tyr Val Cys Met Arg Ile Leu Gly
180 185 190
Val Gly Pro Asp Glu Ser Ala Cys Ala Arg Ala Arg Lys Trp Ile Leu
195 200 205
Asp His Gly Gly Ile Thr Tyr Met Ala Ser Trp Gly Lys Thr Trp Leu
210 215 220
Ser Ile Leu Gly Ile Phe Asp Trp Tyr Gly Cys Asn Pro Met Pro Pro
225 230 235 240
Glu Phe Trp Ile Leu Pro Ser Tyr Leu Pro Ile His Pro Ala Lys Met
245 250 255
Trp Cys Tyr Cys Arg Met Val Tyr Met Pro Met Ser Tyr Leu Tyr Gly
260 265 270
Lys Arg Phe Val Gly Pro Ile Thr Pro Leu Ile Leu Gln Leu Arg Glu
275 280 285
Glu Leu His Thr Gln Pro Tyr His Glu Ile Gln Trp Arg Gln Met Arg
290 295 300
His Arg Cys Gly Lys Glu Asp Leu Tyr Tyr Pro His Ser Leu Ile Gln
305 310 315 320
Asn Phe Leu Trp Asp Gly Leu Tyr Val Ala Ser Glu Pro Leu Leu Thr
325 330 335
Arg Trp Pro Phe Ser Lys Ile Arg Glu Lys Ala Leu Glu Lys Ala Met
340 345 350
Glu His Ile His Tyr Glu Asp Glu Asn Ser Arg Tyr Ile Thr Ile Gly
355 360 365
Cys Val Glu Lys Ala Leu Cys Met Leu Cys Cys Trp Val Glu Asp Pro
370 375 380
Asn Gly Asp Tyr Phe Lys Lys His Leu Ala Arg Ile Pro Asp Tyr Leu
385 390 395 400
Trp Val Ala Glu Asp Gly Met Lys Val Val Ser Phe Gly Ser Gln Leu
405 410 415
Trp Asp Ala Thr Phe Gly Phe Gln Ala Leu Val Ala Ser Asn Leu Thr
420 425 430
Asn Glu Val Ala Pro Thr Leu Val Lys Ala Tyr Asp Phe Val Lys Lys
435 440 445
Cys Gln Val Arg Asn Asn Pro Ser Gly Asp Phe Glu Lys Met Phe Arg
450 455 460
His Ile Ser Lys Gly Ser Trp Thr Phe Ser Asp Gln Asp His Gly Trp
465 470 475 480
Gln Leu Ser Asp Cys Thr Ala Glu Ala Leu Lys Cys Cys Leu Leu Ala
485 490 495
Ala Thr Met Pro Glu Glu Leu Val Gly Glu Lys Leu Asp Pro Gln Trp
500 505 510
Ile Phe Glu Ser Val Asn Ile Ile Leu Ser Leu Gln Glu Pro Lys Thr
515 520 525
Gly Gly Leu Ala Gly Trp Glu Pro Val Arg Ala Gly Gln Trp Met Glu
530 535 540
Met Leu Asn Pro Met Glu Phe Leu Glu Asn Ile Val Ile Glu His Thr
545 550 555 560
Tyr Val Glu Cys Thr Gly Ser
雷公藤三萜合酶TwOSC3及其编码基因与应用专利购买费用说明
![]()
Q:办理专利转让的流程及所需资料
A:专利权人变更需要办理著录项目变更手续,有代理机构的,变更手续应当由代理机构办理。
1:专利变更应当使用专利局统一制作的“著录项目变更申报书”提出。
2:按规定缴纳著录项目变更手续费。
3:同时提交相关证明文件原件。
4:专利权转移的,变更后的专利权人委托新专利代理机构的,应当提交变更后的全体专利申请人签字或者盖章的委托书。
Q:专利著录项目变更费用如何缴交
A:(1)直接到国家知识产权局受理大厅收费窗口缴纳,(2)通过代办处缴纳,(3)通过邮局或者银行汇款,更多缴纳方式
Q:专利转让变更,多久能出结果
A:著录项目变更请求书递交后,一般1-2个月左右就会收到通知,国家知识产权局会下达《转让手续合格通知书》。












动态评分
0.0