








专利摘要
本发明属于生物医药领域,具体涉及TBK1作为E3泛素连接酶的新用途。本发明发现TBK1(TANK结合激酶1)具有泛素化功能,不仅能够在体外发生泛素化,而且能在体内发生泛素化,是一种新的E3泛素连接酶;本发明也发现了TBK1能够降解包括口蹄疫病毒(FMDV)、肠道病毒(EV71)、脑心肌炎病毒(EMCV)以及塞内卡病毒(SVV)在内的小核糖核酸病毒科病毒的结构蛋白VP3,尤其能够特异性地降解FMDV的结构蛋白VP3,可用于制备降解口蹄疫病毒蛋白相关药物,阻止口蹄疫病毒蛋白组装相关药物以及预防或治疗口蹄疫病毒感染相关药物。
权利要求
1.TBK1作为E3泛素连接酶的应用,其特征在于,所述TBK1的核苷酸序列如SEQ ID NO.1所示,所述TBK1的氨基酸序列如SEQ ID NO.2所示。
2.TBK1在制备降解病毒蛋白相关药物中的应用,其特征在于,所述TBK1的核苷酸序列如SEQ ID NO.1所示,所述TBK1的氨基酸序列如SEQ ID NO.2所示。
3.TBK1在制备阻止病毒蛋白组装相关药物中的应用,其特征在于,所述TBK1的核苷酸序列如SEQ ID NO.1所示,所述TBK1的氨基酸序列如SEQ ID NO.2所示。
4.TBK1在制备预防或治疗病毒感染相关药物中的应用,其特征在于,所述TBK1的核苷酸序列如SEQ ID NO.1所示,所述TBK1的氨基酸序列如SEQ ID NO.2所示。
5.如权利要求2-4所述的任一应用,其特征在于,所述的病毒为小核糖核酸病毒科病毒。
6.如权利要求5所述的应用,其特征在于,所述的病毒为FMDV、EV71、EMCV、SVV中的任一种。
7.如权利要求6所述的应用,其特征在于,所述的病毒为FMDV。
8.如权利要求2-3所述的应用,其特征在于,所述的病毒蛋白为结构蛋白。
9.如权利要求8所述的应用,其特征在于,所述的病毒蛋白为VP3。
10.如权利要求2-4所述的任一种应用,其特征在于,所述的TBK1加入药学上可接受的载体和/或辅料,制成粉针剂、胶囊剂、片剂、混悬剂的任一种剂型。
说明书
技术领域
本发明属于生物医药领域,具体涉及TBK1作为E3泛素连接酶的应用。
背景技术
病毒是由一个核酸分子(DNA或RNA)与蛋白质构成的非细胞形态,由一个保护性外壳包裹一段DNA或者RNA,可以利用宿主的细胞系统进行自我复制。病毒可以感染几乎所有具有细胞结构的生命体,可以引发多种疾病。
小核糖核酸(RNA)病毒科是由RNA病毒中最小的类群组成的一科,主要包括肠道病毒属、鼻病毒属、心病毒属以及口疮病毒属。其中口蹄疫属于口疮病毒属,是一种由口蹄疫病毒引起的重要的感染偶蹄动物的疾病,口蹄疫病毒(FMDV)是单股正链RNA病毒,基因组全长约8200bp,属于小RNA病毒科口蹄疫病毒属。FMDV开放阅读框编码一个聚蛋白,翻译后由病毒编码的蛋白酶水解为四个结构蛋白(VP1-VP4)以及八个非结构蛋白(L,2A,2B,2C,3A,3B,3C以及3D)。
目前疫苗接种是特异性预防口蹄疫(FMD)的有效手段,FMD弱毒疫苗和灭活疫苗等常规疫苗都具有良好的免疫原性,在预防和控制FMD的过程中发挥着重要作用。但由于病毒毒力返强、病毒灭活不彻底、活病毒逃逸加工厂等不安全因素,世界上一些地区FMD的暴发似乎与灭活疫苗中残存的活病毒有关。因此,如何更好地抑制病毒蛋白的表达,是预防或控制病毒感染的一种有效途径。
TANK结合激酶1(TBK1)是磷酸化IRF3/7的主要激酶,也是TLR和RLR介导的IFN-β表达中的关键激酶。本发明意外地发现,TBK1不仅能够作为一种新的E3泛素连接酶,而且能有效降解病毒VP3蛋白,进而显著抑制小核糖核酸病毒科,尤其是FMDV的蛋白组装。
发明内容
本发明的目的在于提供一种TBK1作为E3泛素连接酶的应用,所述TBK1的核苷酸序列如SEQ ID NO.1所示,所述TBK1的氨基酸序列如SEQ ID NO.2所示。
本发明的另一目的在于提供一种TBK1在制备降解病毒蛋白相关药物中的应用。
本发明的另一目的在于提供一种TBK1在制备阻止病毒蛋白组装相关药物中的应用。
本发明的另一目的在于提供一种TBK1在制备预防或治疗病毒感染相关药物中的应用。
优选地,所述的病毒为小核糖核酸病毒科病毒。
优选地,所述病毒为FMDV、EV71、EMCV、SVV的任一种病毒。
优选地,所述病毒为FMDV。
优选地,所述的病毒蛋白为结构蛋白。
优选地,所述结构蛋白为VP3。
优选地,所述TBK1加入药学上可接受的载体和/或辅料,制成粉针剂、胶囊剂、片剂、混悬剂的任一种剂型。
本发明的有益效果是:①本发明发现TBK1具有泛素化功能,不仅在体外发生泛素化,还可以在293T细胞中发生体内泛素化,是一种新的E3泛素连接酶;②TBK1能够降解多种小核糖核酸病毒科病毒的结构蛋白VP3,尤其是口蹄疫病毒(FMDV)、肠道病毒(EV71)、脑心肌炎病毒(EMCV)以及塞内卡病毒(SVV),可作为病毒抑制剂应用;③TBK1能够降解多种小核糖核酸病毒科病毒的结构蛋白VP3,而对非结构蛋白3A不具有降解作用,具有特异性。
附图说明
图1 TBK1体外泛素化分析
图2 TBK1体内泛素化分析
图3 TBK1降解VP3蛋白分析
图4 TBK1降解VP3蛋白的剂量依赖性分析
图5 WB检测TBK1-/-MEFs细胞中TBK1的表达情况
图6 FMDV感染TBK1-/-MEFs和TBK1+/+MEFs细胞的分析
图7 TBK1对FMDV、EV71、EMCV、SVV的结构蛋白VP3的影响
图8不同生物体来源的TBK1基因中四个半胱氨酸保守序列比对分析
图9 TBK1基因的四个半光氨酸突变体对VP3蛋白降解的影响
图10突变体TBK1 C426/605A对VP3蛋白降解的影响
图11突变体TBK1C426/605A对天然免疫的影响
图12突变体TBK1C426/605A的泛素化分析
图13 TBK1降解VP3蛋白的特异性分析
具体实施方式
下面通过具体实施例对本发明进行详细的阐述,但本发明的保护范围并不限于以下实施例,任何本领域的技术人员在本发明的基础上,结合本领域公知常识所能想到的技术方案,都属于本发明的保护范围。
在本发明的下述实施例中,所用的实验材料和试剂来源入下:
FMDV由中国农业科学院兰州兽医研究所国家口蹄疫病毒参考实验室提供;
pCDNA3.1-Flag-TBK1质粒、pCDNA3.1-HA-TBK1质粒、pCDNA3.1质粒、MEF细胞、HEK-293T细胞、pCDNA3.1-HA-Ub质粒、pCDNA3.1-HA-VISA质粒、pCDNA3.1-HA-RIG-I质粒、pCDNA3.1-HA-IRF3-5D质粒、pCDNA3.1-HA-IRF7质粒、pCDNA3.1-HA-TBK1质粒、pCDNA3.1-HA-MITA质粒、pCDNA3.1-HA-RIG-I质粒、pCDNA3.1-HA-MDA5质粒、pCDNA3.1-Flag-VP3质粒、pGL-U6-gRNA-TBK1质粒、pST1374-Cas9-D10A质粒、pCDNA3.1-Flag-3A质粒、pCDNA3.1-HA-TBK1C423A质粒、pCDNA3.1-HA-TBK1C426A质粒、pCDNA3.1-HA-TBK1C471A质粒、pCDNA3.1-HA-TBK1C605质粒、pCDNA3.1-HA-TBK1C426/605A质粒、pCDNA3.1-Flag-TBK1C426/605A质粒、pCDNA3.1-FMDV-Myc-VP3质粒、pCDNA3.1-EV71-Myc-VP3质粒、pCDNA3.1-EMCV-Myc-VP3质粒、pCDNA3.1-SVV-Myc-VP3质粒均购买于兰州瑞博莱生物科技有限公司。
实施例1 TBK1作为E3泛素连接酶的活性研究
1.1 TBK1体外泛素化实验
1.1.1质粒的体外转录
(1)根据体外转录试剂盒(TNT Quick Coupled Transcription/TranslationSystems kit)说明,将待检测pCDNA3.1-Flag-TBK1质粒(1μg)和对照质粒pCDNA3.1(1μg)分别用与TNT Quick Master Mix(24μL),Methionine(1.2μL),0.5μg/μL plasmid(2μL),H2O(2.8μL)混合制备成反应物;
(2)将上述反应物置于30℃条件下反应60-90min,获得体外转录产物pCDNA3.1-Flag-TBK1蛋白。
1.1.2泛素化分析实验:
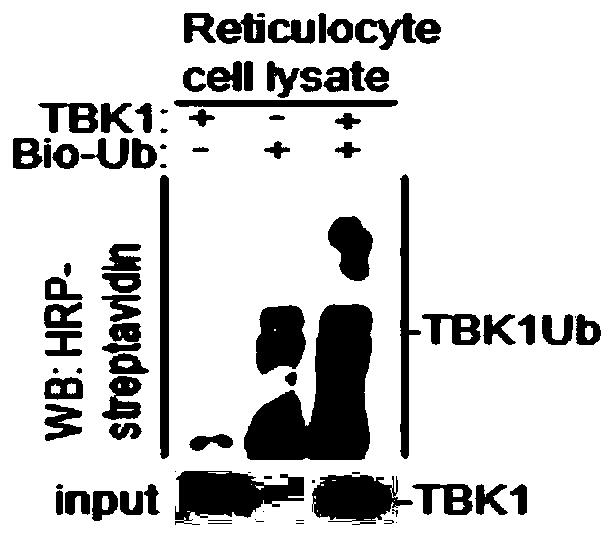
(1)根据体外泛素试剂盒(Enzo Life Sciences)说明书,将上述体外转录产物pCDNA3.1-Flag-TBK1蛋白(7.5μL)与dH2O(14μL),10×Ubiquitionylation Buffer(5μL),100U/ml IPP(10μL),50m M DTT(1μL),0.1M Mg-ATP(2.5μL),Reticulocyte cell lysate(7.5μL),pCDNA3.1-Flag-TBK1,50mM 20×Bio-Ub(2.5μL)均匀混合制备成反应物;
(2)将上述反应物置于37℃下反应30-60min,用HRP-streptavidin进行WesternBlot检测。
1.1.3实验结果
实验结果如图1所示,TBK1与Bio-ub结合,发生了体外泛素化。
1.2TBK1质粒的体内泛素化实验
1.2.1实验步骤
(1)HEK-293T细胞(1×107)铺12孔板,当其密度达到40%-60%时,转染pCDNA3.1-Flag-TBK1(0μg,5μg,10μg)和pCDNA3.1-HA-Ub质粒(2μg);
(2)转染24h后,用100μL含1%SDS的细胞裂解液(20mM Tris、150mM NaCl、10μg/mLleupeptin、1%Triton、10μg/mL aprotinin、1mM EDTA、1mM phenylMethylsulfonylfluoride、pH 7.5)振荡;
(3)震荡结束后在95℃下加热10min,并将900μL不含SDS的裂解液加入其中,4℃12000rpm离心10min,取上清100μL备用,余下上清加入flag抗体与proteinG磁珠。
(4)将样品放在静音混合器4℃上孵育4h。
(5)用细胞裂解液(含0.5M NaCl)洗三次proteinG磁珠,每次1ml,用以洗去非特异性结合的蛋白。然后加上2×SDS聚丙烯酰氨凝胶样品缓冲液60μl,95℃煮5-10min,13000rpm离心1min待检测。
(6)取步骤(3)中备用的100μL上清,加6×SDS聚丙烯酰氨凝胶上样缓冲液20μl,95℃煮5min,用以检测外源性蛋白的表达情况。
1.2.2实验结果
实验结果如图2所示,TBK1与HA-Ub结合,在HEK-293T细胞中发生了泛素化,随着TBK1浓度的增加,检测到的体内泛素化水平逐渐增加,表明TBK1能在体内发生泛素化。
综上实验表明,TBK1具有泛素化功能,是一种新的E3泛素连接酶。
实施例2降解FMDV结构蛋白VP3的信号通路分子分析
2.1实验步骤
(1)HEK-293T细胞铺12孔板,待细胞长至60%-80%时,将信号通路分子的质粒(pCDNA3.1-HA-VISA,pCDNA3.1-HA-RIG-I,pCDNA3.1-HA-IRF3-5D,pCDNA3.1-HA-IRF7,pCDNA3.1-HA-TBK1,pCDNA3.1-HA-MITA,pCDNA3.1-HA-RIG-I,pCDNA3.1-HA-MDA5)各1μg,分别与FMDV的VP3质粒1μg共转染到293T细胞中;
(2)转染24h后收样,用冷却的PBS洗1-2遍,加入100μL的SDS loading buffer进行裂解,WB方法检测VP3蛋白的表达情况。
2.2实验结果
结果如图3所示,只有在TBK1质粒存在的情况下,VP3蛋白没有表达,因此,上述信号通路分子中只有TBK1能够抑制VP3蛋白的表达,说明TBK1能够降解FMDV的VP3蛋白。
实施例3 TBK1降解FMDV结构蛋白VP3的分析
3.1实验步骤
(1)HEK-293T细胞铺12孔板,细胞长至60%-80%时,转染pCDNA3.1-Flag-VP3质粒以及pCDNA3.1-HA-TBK1质粒,其中分别以pCDNA3.1-Flag-VP3质粒1μg(恒量),pCDNA3.1-HA-TBK1质粒为0.5μg、1μg、1.5μg、2μg(变量递增),或者pCDNA3.1-HA-TBK1质粒2μg(恒量),pCDNA3.1-Flag-VP3质粒0.5μg、1μg、1.5μg、2μg(变量递增)进行共转染;
(2)转染24h后收样,用冷却的PBS洗1-2遍,加入100μL的SDS loading buffer裂解,WB检测pCDNA3.1-Flag-VP3质粒以及pCDNA3.1-HA-TBK1质粒的表达情况。
3.2实验结果
结果如图4所示:不管是在pCDNA3.1-Flag-VP3质粒含量不变,pCDNA3.1-HA-TBK1质粒变量递增的情况下,还是pCDNA3.1-HA-TBK1质粒含量不变,pCDNA3.1-Flag-VP3质粒变量递增的情况下,TBK1都能够降解VP3蛋白,并且TBK1的过度表达以剂量依赖性的方式降解了VP3蛋白。
实施例4敲除TBK1基因对VP3蛋白降解的影响
4.1 TBK1-/-MEFs细胞系的构建
4.1.1实验步骤
(1)退火偶联,将浓度为10μM的CRISPR/Cas9 F(CGGCGAGTCAACTCCGGCCA)和R(TGGCCGGAGTTGACTCGCCG)引导序列(5μL),与0.5M NaCl(6μL)以及水(24μL)混合,将退火后的引物置于95℃水浴锅中5min,然后拿出自然降温至室温;
(2)按照说明书,用Fast Digest Bsm BI将pGL-U6-gRNA载体切出粘性末端;将5μL退火后的上述引物与2μL酶切载体用T4连接酶混合,室温连接30min;
(3)取5μL连接产物与50μL感受态DH5α混合,热激30秒,涂板;
(4)挑单克隆,测序,提质粒,得到的重组质粒记为pGL-U6-gRNA-TBK1;
(5)将MEF细胞接种至10cm皿,用8mLDMEM培养基培养过夜(密度约70-80%),并用脂质体2000转染pGL-U6-gRNA-TBK1质粒和pST1374-Cas9-D10A质粒(质粒比为1:1);
(6)转染24h后加入Puromycin培养基筛选,筛选时间为7天;
(7)换为正常的DMEM培养基,从转染后的MEF细胞中获取TBK1基因敲除的MEF细胞系(TBK1-/-MEFs);
(8)通过Western Blot检测TBK1-/-MEFs细胞中TBK1的表达情况。
4.1.2实验结果
结果如图5所示,WB检测表明,TBK1-/-MEFs细胞中,TBK1没有表达,说明TBK1-/-MEFs细胞构建成功。
4.2 FMDV感染TBK1-/-MEFs细胞
4.2.1实验步骤
(1)TBK1+/+MEFs细胞(没有敲除TBK1的野生型MEF细胞)和TBK1-/-MEFs细胞(敲除TBK1的MEF细胞)铺于12孔板,12h后接FMDV(MOI=0.1),分别在12h和16h收集样品,进行绝对定量Q-PCR实验,检测FMDV基因的拷贝数。
(2)在步骤(1)的样品中加入TRIZOL试剂(500μL)裂解5min,再加入氯仿(100μL)混匀,室温放置10min后,在4℃,12000rpm离心15min,取上清;
(3)取步骤(2)中的上清200μL到1.5ml EP管中,并加入异丙醇(200μL)混匀,室温放置10min,4℃,12000rpm离心10min;
(4)去上清后加入1ml 75%乙醇,4℃12000rpm离心5min,去上清干燥并加入DEPC水溶解。
4.2.2 Q-PCR反应体系
反应试剂:2×one step RT-PCR bufferⅢ(12.5ml),Takara Ex Taq HS(0.5ml),Primer Scrip RT Enzyme mixⅡ(0.5μL),正向引物(5’ACTGGGTTTTACAAACCTGTGA-3C,0.5μL),反向引物(5’GCGAGTCCTGCCACGGA-3C,0.5μL),探针(5FAM-TCCTTTGCACGCCGTGGGAC-TAMRA--3A,1μL),RNA(2μL),H2O(7.5μL);
反应条件:42℃,15min;95℃,10s;55℃,30s;72℃,30s;2-4共40个循环。
4.2.3实验结果
实验结果如图6所示,不管是在FMDV感染MEF细胞后12h还是16h后,TBK1-/-MEFs细胞中FMDV基因拷贝数均显著高于TBK1+/+MEFs细胞,由此说明,敲除TBK1基因后,FMDV感染MEF细胞的能力显著增强。
实施例5 TBK1对其它小RNA病毒科VP3蛋白降解
5.1实验步骤
(1)293T细胞铺于12孔板,12h后分别共转染pCDNA3.1载体上带HA标签的TBK1质粒(pCDNA3.1-HA-TBK1,0μg,0.25μg,0.5μg)和pCDNA3.1载体上带Myc标签的FMDV、EV71、EMCV以及SVV的VP3结构蛋白质粒(pCDNA3.1-FMDV-Myc-VP3,pCDNA3.1-EV71-Myc-VP3,pCDNA3.1-EMCV-Myc-VP3和pCDNA3.1-SVV-Myc-VP3,各1μg);
(2)转染24h后,用SDS-loading buffer裂解,WB检测TBK1和VP3的表达。
5.2实验结果
结果如图7所示,随着TBK1含量的增加,FMDV、EV71、EMCV、SVV结构蛋白VP3的表达显著降低或者抑制,说明TBK1对小核糖核酸病毒科(FMDV、EV71、EMCV、SVV)的结构蛋白VP3均有降解作用。
实施例6 TBK1降解VP3蛋白的泛素化位点分析
6.1 TBK1中四种保守半胱氨酸的序列分析
将人、小鼠、牛、猴、猪、大鼠六种不同生物体来源的TBK1半胱氨酸保守序列进行比对,分析鉴定了TBK1中的四个保守半胱氨酸C423、C426、C471和C605,其序列如图8所示。
6.2 TBK1降解VP3蛋白的位点分析
(1)将上述四种半胱氨酸C423、C426、C471和C605分别突变为丙氨酸,构建TBK1突变体(TBK1C423A、TBK1C426A、TBK1C471A、TBK1C605,由兰州瑞博莱生物科技有限公司构建),并获得pCDNA3.1-HA-TBK1C423A质粒、pCDNA3.1-HA-TBK1C426A质粒、pCDNA3.1-HA-TBK1C471A质粒、pCDNA3.1-HA-TBK1C605质粒,通过转染实验,检测TBK1四种突变体对FMDV结构蛋白VP3的影响。
结果如图9所示,将C423、和C471位点处的半胱氨酸突变为丙氨酸时产生的TBK1突变体(TBK1C423A和TBK1C471A)仍然能够降解FMDV的VP3结构蛋白,说明C423、和C471并不是降解FMDV的VP3结构蛋白的位点;而将C426和C605突变为丙氨酸时产生TBK1突变体(TBK1C426A和TBK1C605)后,不会降解FMDV结构蛋白VP3,说明C426和C605是降解FMDV结构蛋白VP3的位点。
(2)将C426和C605半胱氨酸同时突变为丙氨酸,得到突变体TBK1C426/605A,并构建pCDNA3.1-HA-TBK1C426/605A质粒,检测TBK1C426/605A突变体对FMDV结构蛋白VP3降解的影响。
实验结果如图10显示,将C426和C605位点处的半胱氨酸同时突变为丙氨酸,得到突变体TBK1C426/605A对FMDV结构蛋白VP3的表达没有影响。
上述实验表明C426和C605位点处的半胱氨酸分别或同时突变为丙氨酸后,都不会再降解FMDV的结构蛋白VP3,再次证明了C426和C605是TBK1降解FMDV的结构蛋白VP3的关键位点。
6.3 TBK1 C426/605A突变体对其天然免疫活性的影响
将TBK1的C426和C605半胱氨酸同时突变为丙氨酸,以评估其在TBK1激活中的作用。
结果如图11所示,突变体TBK1C426/605A对TBK1介导的IRF3磷酸化和TBK1的自磷酸化没有影响。说明,TBK1通过C426和C605位点降解VP3蛋过程与其天然免疫活性无关。
6.4 TBK1 C426/605A突变体体外泛素化分析
(1)根据实施例1中质粒的体外转录操作过程,将pCDNA3.1-Flag-TBK1、pCDNA3.1-Flag-TBK1C426/605A、以及空载pCDNA3.1质粒各1μg,进行体外转录。
(2)根据实施例1中的体外泛素化分析步骤,对步骤(1)中体外转录的产物进行体外泛素化分析。
(3)实验结果如图12所示,将TBK1的C426和C605位点的半胱氨酸同时突变为丙氨酸得到的突变体TBK1C426/605A的体外泛素化现象消失。说明,TBK1的C426和C605位点是通过泛素化作用来降解FMDV结构蛋白VP3的关键位点。
实施例7 TBK1降解FMDV的VP3蛋白的特异性分析
7.1实验步骤
(1)293T细胞铺12孔板,细胞长至60%-80%时,TBK1质粒(0.25μg)分别共转染pCDNA3.1-Flag-VP3质粒(1μg)和pCDNA3.1-Flag-3A质粒(1μg);
(2)转染24h后收样,用冷却的PBS洗1-2遍,加入SDS loading buffer(100μL)裂解;
(3)WB检测TBK1质粒,VP3蛋白以及3A质粒的表达情况。
7.2实验结果
结果如图13所示,TBK1质粒能够降解FMDV的结构蛋白VP3,但对非结构蛋白3A无影响。说明TBK1质粒能够特异性降解FMDV的结构蛋白VP3。
序列表
<110> 中国农业科学院兰州兽医研究所
<120> TBK1作为E3泛素连接酶的应用
<160> 2
<170> SIPOSequenceListing 1.0
<210> 1
<211> 2190
<212> DNA
<213> 人(Homo sapiens)
<400> 1
atgcagagca cttctaatca tctgtggctt ttatctgata ttttaggcca aggagctact 60
gcaaatgtct ttcgtggaag acataagaaa actggtgatt tatttgctat caaagtattt 120
aataacataa gcttccttcg tccagtggat gttcaaatga gagaatttga agtgttgaaa 180
aaactcaatc acaaaaatat tgtcaaatta tttgctattg aagaggagac aacaacaaga 240
cataaagtac ttattatgga attttgtcca tgtgggagtt tatacactgt tttagaagaa 300
ccttctaatg cctatggact accagaatct gaattcttaa ttgttttgcg agatgtggtg 360
ggtggaatga atcatctacg agagaatggt atagtgcacc gtgatatcaa gccaggaaat 420
atcatgcgtg ttatagggga agatggacag tctgtgtaca aactcacaga ttttggtgca 480
gctagagaat tagaagatga tgagcagttt gtttctctgt atggcacaga agaatatttg 540
caccctgata tgtatgagag agcagtgcta agaaaagatc atcagaagaa atatggagca 600
acagttgatc tttggagcat tggggtaaca ttttaccatg cagctactgg atcactgcca 660
tttagaccct ttgaagggcc tcgtaggaat aaagaagtga tgtataaaat aattacagga 720
aagccttctg gtgcaatatc tggagtacag aaagcagaaa atggaccaat tgactggagt 780
ggagacatgc ctgtttcttg cagtctttct cggggtcttc aggttctact tacccctgtt 840
cttgcaaaca tccttgaagc agatcaggaa aagtgttggg gttttgacca gttttttgca 900
gaaactagtg atatacttca ccgaatggta attcatgttt tttcgctaca acaaatgaca 960
gctcataaga tttatattca tagctataat actgctacta tatttcatga actggtatat 1020
aaacaaacca aaattatttc ttcaaatcaa gaacttatct acgaagggcg acgcttagtc 1080
ttagaacctg gaaggctggc acaacatttc cctaaaacta ctgaggaaaa ccctatattt 1140
gtagtaagcc gggaacctct gaataccata ggattaatat atgaaaaaat ttccctccct 1200
aaagtacatc cacgttatga tttagacggg gatgctagca tggctaaggc aataacaggg 1260
gttgtgtgtt atgcctgcag aattgccagt accttactgc tttatcagga attaatgcga 1320
aaggggatac gatggctgat tgaattaatt aaagatgatt acaatgaaac tgttcacaaa 1380
aagacagaag ttgtgatcac attggatttc tgtatcagaa acattgaaaa aactgtgaaa 1440
gtatatgaaa agttgatgaa gatcaacctg gaagcggcag agttaggtga aatttcagac 1500
atacacacca aattgttgag actttccagt tctcagggaa caatagaaac cagtcttcag 1560
gatatcgaca gcagattatc tccaggtgga tcactggcag acgcatgggc acatcaagaa 1620
ggcactcatc cgaaagacag aaatgtagaa aaactacaag tcctgttaaa ttgcatgaca 1680
gagatttact atcagttcaa aaaagacaaa gcagaacgta gattagctta taatgaagaa 1740
caaatccaca aatttgataa gcaaaaactg tattaccatg ccacaaaagc tatgacgcac 1800
tttacagatg aatgtgttaa aaagtatgag gcatttttga ataagtcaga agaatggata 1860
agaaagatgc ttcatcttag gaaacagtta ttatcgctga ctaatcagtg ttttgatatt 1920
gaagaagaag tatcaaaata tcaagaatat actaatgagt tacaagaaac tctgcctcag 1980
aaaatgttta cagcttccag tggaatcaaa cataccatga ccccaattta tccaagttct 2040
aacacattag tagaaatgac tcttggtatg aagaaattaa aggaagagat ggaaggggtg 2100
gttaaagaac ttgctgaaaa taaccacatt ttagaaaggt ttggctcttt aaccatggat 2160
ggtggccttc gcaacgttga ctgtctttag 2190
<210> 2
<211> 729
<212> PRT
<213> 人(Homo sapiens)
<400> 2
Met Gln Ser Thr Ser Asn His Leu Trp Leu Leu Ser Asp Ile Leu Gly
1 5 10 15
Gln Gly Ala Thr Ala Asn Val Phe Arg Gly Arg His Lys Lys Thr Gly
20 25 30
Asp Leu Phe Ala Ile Lys Val Phe Asn Asn Ile Ser Phe Leu Arg Pro
35 40 45
Val Asp Val Gln Met Arg Glu Phe Glu Val Leu Lys Lys Leu Asn His
50 55 60
Lys Asn Ile Val Lys Leu Phe Ala Ile Glu Glu Glu Thr Thr Thr Arg
65 70 75 80
His Lys Val Leu Ile Met Glu Phe Cys Pro Cys Gly Ser Leu Tyr Thr
85 90 95
Val Leu Glu Glu Pro Ser Asn Ala Tyr Gly Leu Pro Glu Ser Glu Phe
100 105 110
Leu Ile Val Leu Arg Asp Val Val Gly Gly Met Asn His Leu Arg Glu
115 120 125
Asn Gly Ile Val His Arg Asp Ile Lys Pro Gly Asn Ile Met Arg Val
130 135 140
Ile Gly Glu Asp Gly Gln Ser Val Tyr Lys Leu Thr Asp Phe Gly Ala
145 150 155 160
Ala Arg Glu Leu Glu Asp Asp Glu Gln Phe Val Ser Leu Tyr Gly Thr
165 170 175
Glu Glu Tyr Leu His Pro Asp Met Tyr Glu Arg Ala Val Leu Arg Lys
180 185 190
Asp His Gln Lys Lys Tyr Gly Ala Thr Val Asp Leu Trp Ser Ile Gly
195 200 205
Val Thr Phe Tyr His Ala Ala Thr Gly Ser Leu Pro Phe Arg Pro Phe
210 215 220
Glu Gly Pro Arg Arg Asn Lys Glu Val Met Tyr Lys Ile Ile Thr Gly
225 230 235 240
Lys Pro Ser Gly Ala Ile Ser Gly Val Gln Lys Ala Glu Asn Gly Pro
245 250 255
Ile Asp Trp Ser Gly Asp Met Pro Val Ser Cys Ser Leu Ser Arg Gly
260 265 270
Leu Gln Val Leu Leu Thr Pro Val Leu Ala Asn Ile Leu Glu Ala Asp
275 280 285
Gln Glu Lys Cys Trp Gly Phe Asp Gln Phe Phe Ala Glu Thr Ser Asp
290 295 300
Ile Leu His Arg Met Val Ile His Val Phe Ser Leu Gln Gln Met Thr
305 310 315 320
Ala His Lys Ile Tyr Ile His Ser Tyr Asn Thr Ala Thr Ile Phe His
325 330 335
Glu Leu Val Tyr Lys Gln Thr Lys Ile Ile Ser Ser Asn Gln Glu Leu
340 345 350
Ile Tyr Glu Gly Arg Arg Leu Val Leu Glu Pro Gly Arg Leu Ala Gln
355 360 365
His Phe Pro Lys Thr Thr Glu Glu Asn Pro Ile Phe Val Val Ser Arg
370 375 380
Glu Pro Leu Asn Thr Ile Gly Leu Ile Tyr Glu Lys Ile Ser Leu Pro
385 390 395 400
Lys Val His Pro Arg Tyr Asp Leu Asp Gly Asp Ala Ser Met Ala Lys
405 410 415
Ala Ile Thr Gly Val Val Cys Tyr Ala Cys Arg Ile Ala Ser Thr Leu
420 425 430
Leu Leu Tyr Gln Glu Leu Met Arg Lys Gly Ile Arg Trp Leu Ile Glu
435 440 445
Leu Ile Lys Asp Asp Tyr Asn Glu Thr Val His Lys Lys Thr Glu Val
450 455 460
Val Ile Thr Leu Asp Phe Cys Ile Arg Asn Ile Glu Lys Thr Val Lys
465 470 475 480
Val Tyr Glu Lys Leu Met Lys Ile Asn Leu Glu Ala Ala Glu Leu Gly
485 490 495
Glu Ile Ser Asp Ile His Thr Lys Leu Leu Arg Leu Ser Ser Ser Gln
500 505 510
Gly Thr Ile Glu Thr Ser Leu Gln Asp Ile Asp Ser Arg Leu Ser Pro
515 520 525
Gly Gly Ser Leu Ala Asp Ala Trp Ala His Gln Glu Gly Thr His Pro
530 535 540
Lys Asp Arg Asn Val Glu Lys Leu Gln Val Leu Leu Asn Cys Met Thr
545 550 555 560
Glu Ile Tyr Tyr Gln Phe Lys Lys Asp Lys Ala Glu Arg Arg Leu Ala
565 570 575
Tyr Asn Glu Glu Gln Ile His Lys Phe Asp Lys Gln Lys Leu Tyr Tyr
580 585 590
His Ala Thr Lys Ala Met Thr His Phe Thr Asp Glu Cys Val Lys Lys
595 600 605
Tyr Glu Ala Phe Leu Asn Lys Ser Glu Glu Trp Ile Arg Lys Met Leu
610 615 620
His Leu Arg Lys Gln Leu Leu Ser Leu Thr Asn Gln Cys Phe Asp Ile
625 630 635 640
Glu Glu Glu Val Ser Lys Tyr Gln Glu Tyr Thr Asn Glu Leu Gln Glu
645 650 655
Thr Leu Pro Gln Lys Met Phe Thr Ala Ser Ser Gly Ile Lys His Thr
660 665 670
Met Thr Pro Ile Tyr Pro Ser Ser Asn Thr Leu Val Glu Met Thr Leu
675 680 685
Gly Met Lys Lys Leu Lys Glu Glu Met Glu Gly Val Val Lys Glu Leu
690 695 700
Ala Glu Asn Asn His Ile Leu Glu Arg Phe Gly Ser Leu Thr Met Asp
705 710 715 720
Gly Gly Leu Arg Asn Val Asp Cys Leu
725
TBK1作为E3泛素连接酶的应用专利购买费用说明
![]()
Q:办理专利转让的流程及所需资料
A:专利权人变更需要办理著录项目变更手续,有代理机构的,变更手续应当由代理机构办理。
1:专利变更应当使用专利局统一制作的“著录项目变更申报书”提出。
2:按规定缴纳著录项目变更手续费。
3:同时提交相关证明文件原件。
4:专利权转移的,变更后的专利权人委托新专利代理机构的,应当提交变更后的全体专利申请人签字或者盖章的委托书。
Q:专利著录项目变更费用如何缴交
A:(1)直接到国家知识产权局受理大厅收费窗口缴纳,(2)通过代办处缴纳,(3)通过邮局或者银行汇款,更多缴纳方式
Q:专利转让变更,多久能出结果
A:著录项目变更请求书递交后,一般1-2个月左右就会收到通知,国家知识产权局会下达《转让手续合格通知书》。












动态评分
0.0