

IPC分类号 : C12N9/04I,C12N15/53I,C12N15/81I,C12Q1/26I,C12Q1/00I,H01M8/16I







专利摘要
本发明属于生物技术领域,具体涉及一种葡萄糖脱氢酶突变体及其制备方法。其中,所述葡萄糖脱氢酶来源于土曲霉,所述突变体是通过将所述葡萄糖脱氢酶的第406位氨基酸置换成其它氨基酸得到的。本发明制备得到的FAD‑GDH突变体与野生型FAD‑GDH相比,活性和底物专一性都得到了非常大程度的提高。
权利要求
1.一种葡萄糖脱氢酶的突变体,其特征在于,所述葡萄糖脱氢酶来源于土曲霉,所述突变体是通过所述葡萄糖脱氢酶的第406位氨基酸置换成缬氨酸得到的;所述葡萄糖脱氢酶的氨基酸序列如SEQ ID NO:1所示,核苷酸序列如SEQ ID NO:2所示。
2.一种核苷酸序列,其特征在于,所述核苷酸序列编码如权利要求1所述的葡萄糖脱氢酶的突变体。
3.一种重组载体,其特征在于,所述重组载体包括如权利要求2所述的核苷酸序列。
4.一种使用如权利要求2所述的核苷酸序列或如权利要求3所述的重组载体转化的转化体。
5.一种如权利要求1所述的葡萄糖脱氢酶的突变体的制备方法,其特征在于,包括以下步骤:
1)通过PCR扩增SEQ ID NO:2序列,并与载体pPICK9连接,得到重组质粒pPICK9-fad-gdh;
2)以所述重组质粒pPICK9-fad-gdh为模板,Y406X-F和Y406X-R为引物,进行饱和突变,得到突变质粒;
3)将所述突变质粒进行线性化处理,并转化入宿主细胞中;
4)培养步骤3)得到的细胞,收集培养基上清,分离纯化得到所述葡萄糖脱氢酶的突变体;
Y406X-F和Y406X-R引物序列分别为SEQ ID NO:5和SEQ ID NO:6。
6.一种用于葡萄糖测定的组合物,其特征在于,所述组合物包括如权利要求1所述的葡萄糖脱氢酶的突变体。
7.根据权利要求1所述的葡萄糖脱氢酶的突变体在制备血糖试纸、葡萄糖传感器或生物燃料电池方面的用途。
说明书
技术领域
本发明属于生物技术领域,具体涉及一种葡萄糖脱氢酶突变体及其制备方法。
背景技术
葡萄糖脱氢酶(Glucose dehydrogenase,GDH)是一类能催化葡萄糖转化为葡萄糖酸内酯的酶。该反应中葡萄糖失去电子,传递给了GDH的辅酶或辅基。在其它电子受体存在的情况下,电子可以继续进行传递,以实现GDH的再生。因此该酶在血糖检验试纸、生物燃料电池和电化学生物传感器等方面都有广泛的应用。GDH作为以上设备的一个重要的生物元件,在很大程度上决定了设备的性能。因此,酶学性质优良的GDH的获得对相关生物医学工程设备的研发来说至关重要。
对于糖尿病患者来说日常的血糖检测工作非常重要。伴随着血糖监测技术的快速发展,监测手段越来越方便、快速和准确,便携式血糖监测系统已经广泛应用于病人自我监测和医院内即时检验(point-of-care testing,POCT)的血糖监测中。血糖检测用酶作为便携式血糖监测系统的核心部分,其性能对检测结果的准确性起到至关重要的作用。目前血糖检测用酶有葡萄糖氧化酶(Glucose oxidase,GOD)、己糖激酶(Hexokinase,HK)和GDH等多种。
最早采用的是GOD,其血糖检测原理是利用消耗氧气催化葡萄糖氧化,反应依赖氧气的特点使该检测方法受空气中氧分压的影响较大,往往会产生较大的测量误差。
另一种检测方法是HK法,但该法需要与葡萄糖-6-磷酸脱氢酶联用,需要消耗NADP,当样品中存在能消耗NADP的成分时,会对检测结果造成干扰。另外,由于试剂稳定性较差、价格昂贵,因而限制了HK法在临床上的普及。
GDH是一种氧化还原酶,在该酶的催化下,葡萄糖发生脱氢氧化反应,生成葡萄糖酸内酯。在该反应过程中,生成的还原型辅基或辅酶的含量与葡萄糖的浓度成正相关,且一般浓度的抗凝剂、防腐剂、尿酸、胆红素等对血糖测定不产生干扰。相对于传统的GOD法和HK法而言,GDH法操作更简便,只需一种酶参与反应即可实现,并且既可以进行连续监测(λ=340nm)又可以进行终点分析,因此有利于构建起方便快捷且自动化分析模式。
GDH根据所结合辅基或辅酶的不同,可以分为PQQ依赖的GDH(PQQ-GDH)、NAD依赖的GDH(NAD-GDH)、和FAD依赖的GDH(FAD-GDH)三种。可溶表达的sPQQ-GDH底物专一性很差,底物谱太宽;膜结合型的mPQQ-GDH系膜蛋白,表达和分离纯化都很困难,这些都严重限制了其在血糖测定方面的应用。NAD-GDH由于辅基结合不牢固,需要不断添加辅基来维持其活性,也不利于血糖检测。而FAD-GDH辅基结合紧密不易脱落,其次反应过程中不需要氧气的参与,因此不管是检测的准确性还是保存的长久性都优于葡萄糖氧化酶和其他辅基依赖性GDH,并且市面主流的血糖仪的使用的核心酶也是FAD-依赖的葡萄糖脱氢酶,一般自开封起可使用到有效期结束。
但野生型FAD-GDH对麦芽糖和木糖仍有一定的脱氢氧化活性,例如对麦芽糖的活性约为葡萄糖的20%,对木糖的活性约为10%。因此,野生型FAD-GDH对木糖和麦芽糖会有轻微的反应,在检测血糖时会出现假性血糖。
发明内容
为了解决上述技术问题,本发明提供了一种对麦芽糖和木糖没有活性的新型葡萄糖脱氢酶突变体,及其制备方法。
因此,本发明的目的之一在于提供一种葡萄糖脱氢酶突变体。
本发明的另一目的在于提供一种编码上述突变体的核苷酸序列。
本发明的又一目的在于提供一种包含上述基因的重组载体。
本发明的又一目的在于提供一种包含上述重组载体的转化体。
本发明的又一目的在于提供一种上述葡萄糖脱氢酶突变体的制备方法。
本发明的又一目的在于提供一种包含上述葡萄糖脱氢酶突变体的用于葡萄糖测定的组合物。
本发明的又一目的在于提供上述突变体在制备血糖试纸、葡萄糖传感器或生物燃料电池方面的用途。
本发明所采用的技术方案如下文所述。
本发明的葡萄糖脱氢酶的突变体,所述葡萄糖脱氢酶来源于土曲霉,所述突变体是通过所述葡萄糖脱氢酶的第406位氨基酸置换成其它氨基酸得到的。
进一步地,所述第406位氨基酸为酪氨酸,所述其它氨基酸为除酪氨酸以外的其它天然氨基酸。
进一步地,所述第406位氨基酸被置换成缬氨酸、赖氨酸、谷氨酸、亮氨酸、天冬氨酸、精氨酸或苏氨酸,优选被置换成缬氨酸、赖氨酸或精氨酸,最有选被置换成缬氨酸。
进一步地,所述葡萄糖脱氢酶的氨基酸序列如SEQ ID NO:1所示,核苷酸序列如SEQ ID NO:2所示。
本发明还提供一种核苷酸序列,所述核苷酸序列编码如上所述的葡萄糖脱氢酶的突变体。
本发明还提供一种包含上述核苷酸序列的重组载体。
进一步地,所述载体为质粒pPICK9。
本发明还提供一种使用如上所述的核苷酸序列或如上所述的重组载体转化的转化体。
本发明还提供一种上述葡萄糖脱氢酶突变体的制备方法,包括以下步骤:
1)通过PCR扩增SEQ ID NO:2序列,并与载体pPICK9连接,得到充足质粒pPICK9-fad-gdh;
2)以所述重组质粒pPICK9-fad-gdh为模板,Y406X-F/R为引物,进行饱和突变,得到突变质粒;
3)将所述突变质粒进行线性化处理,并转化入宿主细胞中;
4)培养步骤3)得到的细胞,收集培养基上清,分离纯化得到所述葡萄糖脱氢酶突变体。
其中,所述Y406X是指用氨基酸X将第406位的氨基酸Y替换,Y表示第406位氨基酸为酪氨酸,X表示除酪氨酸外的其它天然氨基酸。
进一步地,步骤1)中,所述PCR扩增的引物序列为SEQ ID NO:3和SEQ ID NO:4.
进一步地,步骤2)中,所述Y406X-F/R引物序列为SEQ ID NO:5和SEQ ID NO:6。
进一步地,步骤3)中,所述宿主细胞为毕赤酵母细胞。
进一步地,步骤3)中,还包括将转化后的宿主细胞进行筛选的步骤。
进一步地,采用含有遗传霉素的YPD培养基对所述转化后的宿主细胞进行筛选。
进一步地,步骤4)中,可以采用异丙醇进行纯化。
本发明还提供一种用于葡萄糖测定的组合物,所述组合物包括如上所述的葡萄糖脱氢酶的突变体。
本发明还提供一种上述葡萄糖突突变体在制备血糖试纸、葡萄糖传感器或生物燃料电池方面的用途。
本发明对来源于土曲霉的FAD-GDH进行了相关的融合表达并对氨基酸残基进行分析和突变等工作,明确了Y406位点的改变对整个酶的底物专一性有较大的影响。采用PCR定点饱和突变技术对Y406位点进行改造,突变后的FAD-GDH与野生型FAD-GDH相比,底物专一性都得到了非常大程度的提高。
附图说明
图1示出了使用遗传霉素对多拷贝重组菌的筛选结果;其中,A为电转后,MD平板长出的白色单菌落;B为培养前接种含0.5mg/mL遗传霉素的YPD平板;C为含0.5mg/mL遗传霉素的YPD平板长出的白色单菌落;D为含1.0mg/mL遗传霉素的YPD平板长出的白色单菌落。
图2示出了葡萄糖脱氢酶突变体的SDS-PAGE蛋白电泳图;
图3示出了葡萄糖脱氢酶突变体的SDS-PAGE蛋白电泳图;
图4示出了葡萄糖脱氢酶突变体的SDS-PAGE蛋白电泳图;
图5示出了葡萄糖脱氢酶突变体Y406V与野生型FAD-GDH蛋白纯化的SDS-PAGE分析结果;
图6示出了葡萄糖脱氢酶突变体Y406V和野生型FAD-GDH的酶活和pH值关系图;
图7示出了葡萄糖脱氢酶突变体Y406V和野生型FAD-GDH的酶活和温度关系图;
图8示出了葡萄糖脱氢酶突变体Y406V和野生型FAD-GDH的pH值稳定性图;
图9示出了葡萄糖脱氢酶突变体Y406V和野生型FAD-GDH的热稳定性图。
具体实施方式
以下结合附图和具体的实施例对本发明的技术方案做进一步说明,但本发明并不限于这些具体实施方式。实施例中所用的材料、试剂等,如无特殊说明,均可从商业途径得到。
实施例1.重组质粒pPIC9K-fad-gdh的构建
利用无限制(restriction-free,RF)克隆法,以9K99a F和9K99a R作为引物,fad-gdh(SEQ ID NO:2)作为模板,进行PCR扩增。使用SanPrep柱式PCR产物纯化试剂盒(生工生物工程(上海)有限公司),将PCR扩增产物进行纯化,纯化后的产物作为引物,以pPIC9K质粒为模板,使用Takara PrimeSTAR GXL DNA聚合酶进行第二次PCR,得到重组质粒pPIC9K-fad-gdh。
其中,引物序列为:
9K99a F:
5-CTCTCGAGAAAAGAGAGGCTGAAGCTTCACACAGGAAACAGACCATG-3′(SEQ ID NO:3),
9K99a R:
5-GGAACAGTCATGTCTAAGGCGAATTACATCCGCCAAAACAGCCAAGC-3′(SEQ ID NO:4)。
实施例2.得到fad-gdh的Y406位点的突变构建体
使用Y406位点的随机突变引物Y406X-F/R,以实施例1制备得到的重组质粒pPIC9K-fad-gdh为模板进行定点饱和突变。将饱和突变PCR产物转化到XL10-GOLD感受态细胞中进行克隆。使用提取试剂盒(生工生物工程(上海)有限公司)提取质粒,并对质粒进行测序。从中筛选得到19种在FAD-GDH的Y406位点突变的质粒pPIC9K-fad-gdh。
其中,随机突变引物为:
Y406X-F:
5′-TGCCGAAGTTTTAAACNNNCCAGGCTCCGCCAC-3′(SEQ ID NO:5)
Y406X-R:
5′-GTGGCGGAGCCTGGNNNGTTTAAAACTTCGGCA-3′(SEQ ID NO:6)。
实施例3.FAD-GDH在重组毕赤酵母菌株中的表达
将实施例2制备得到突变质粒进行线性化处理,后使用基因导入仪突变质粒转入毕赤酵母细胞中。
将重组毕赤酵母涂于MD平板,30℃恒温培养2~3天至白色菌落长出。用牙签挑取MD平板上的白色单菌落,接种于含0.5mg/mL遗传霉素的YPD平板中,30℃培养2~3天至白色菌落长出。用牙签挑取0.5mg/mL遗传霉素的YPD平板上生长的白色单菌落,接种至含1.0mg/mL遗传霉素的YPD平板中,30℃培养2~3天至白色菌落。以Y406E(第406位酪氨酸被谷氨酸替代)为例,结果如图1A~1D所示,图1示出了使用遗传霉素对多拷贝重组菌的筛选结果。其中,A为电转后,MD平板长出的白色单菌落;B为培养前接种含0.5mg/mL遗传霉素的YPD平板;C为含0.5mg/mL遗传霉素的YPD平板长出的白色单菌落;D为含1.0mg/mL遗传霉素的YPD平板长出的白色单菌落。
挑取含1.0mg/mL遗传霉素的YPD平板上生长的圆润白色单菌落于50mL BMGY培养基中,30℃、250rpm培养至A600为6.0。
取600μL上述菌液进行保菌处理,剩余菌液用50mL离心管5000rpm、4℃离心5min收集菌体,用新鲜的BMMY培养基重悬菌体,30℃、250rpm培养。每隔24h取样测定菌液浓度和样品上清的酶活力,同时加入甲醇至终浓度为1.5%,至酶活不再升高时停止诱导。
实施例4.经重组毕赤酵母表达的FAD-GDH蛋白的初步纯化
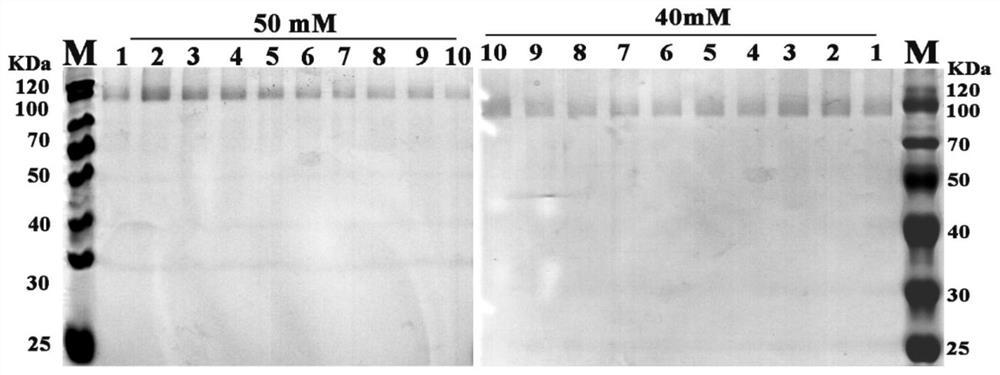
将实施例3制备得到的菌株的菌液8000×g、4℃离心10min取上清。加入预冷的异丙醇至终浓度为40%,马上4℃、12000×g、离心15min,立刻弃上清。所得沉淀用pH 7.4的MOPS进行溶解,得到纯化的蛋白。获得的蛋白经SDS-PAGE蛋白电泳进行检测。SDS-PAGE蛋白电泳结果如图2~4所示。
图2中的9条泳道分别为:M:Marker;1:野生型FAD-GDH(WT);2:Y406M;3:Y406G;4:Y406F;5:Y406T;6:Y406P;7:Y406K;8:Y406Q。
图3中的8条泳道分别为:M:Marker;1:Y406C;2:Y406I;3:Y406V;4:Y406A;5:Y406L;6:Y406E;7:WT。
图4中7条泳道分别为:M:Marker;1:Y406W;2:Y406S;3:Y406N;4:Y406D;5:Y406R;6:Y406H。
从图中可以看出蛋白条带呈现拖尾状在66.2KDa和116KDa之间,与目的蛋白分子量大小一致。
实施例5.对获得的FAD-GDH蛋白的酶活力的测定
FAD-GDH氧化葡萄糖的酶活力测定原理如下:
D-Glucose+ox-PMS→D-glucono-δ-lactone+re-PMS
re-PMS+ox-DCIP→ox-PMS+re-DCP
DCIP(2,6-Dichlorophenolindophenol sodium salt,2,6-二氯靛酚钠)为氧化态的时候溶液为蓝色,600nm波长下具有吸收波长,还原态的DCIP溶液接近无色。若发生上述反应,蓝色溶液会逐渐褪色且吸光值也会相应地减少,因此通过A600值的变化我们可以对酶活力进行测定和计算。反应体系及其浓度如表1。
表1.葡萄糖脱氢酶的酶活力测定体系
PMS:Phenazine methosulfate,吩嗪硫酸甲酯。
表1中的MOPS(3-morpholinopropane sulfonic acid,3-吗啉基丙磺酸)的pH值为7.4,反应温度为25℃,测定其吸光值的变化(ΔABs/min),DCIP的摩尔吸光系数为16.3×10
酶活性(U/mL)=-ΔABs/16.3×2.0/0.01×N(酶的稀释倍数)
取实施例4初步纯化后的蛋白,按照上述酶活力测定方法,将反应体系的底物分别换成1M D-麦芽糖,D-木糖,D-半乳糖,L-阿拉伯糖,海藻糖,D-果糖,D-山梨醇和D-甘露糖,测定酶活力,并与以D-葡萄糖为底物时测定的酶活力进行比较。每个样品做三个平行样。结果如表2所示。
表2野生型FAD-GDH和突变体底物专一性比较
注:WT:野生型FAD-GDH;ND
从表2所示的结果可以看出,Y460V、Y406K、Y406R三种突变体底物专一性相比其它突变体更高,选择其中底物专一性最高的Y406V进行后续实验。
实施例6经重组毕赤酵母表达的FAD-GDH蛋白的精细纯化
对实施例4初步纯化后的野生型FAD-GDH及其突变体进行精细纯化,步骤如下:
(1)配制纯化缓冲液Buffer A:20mmol/L Tris,0.5mol/L NaCl,20mmol/L,pH 8.0的咪唑,超声除气后备用;
(2)配制纯化缓冲液Buffer B:20mmol/L Tris,0.5mol/L NaCl,250mmol/L,pH8.0的咪唑,超声除气后备用;
(3)组装完蠕动泵之后将镍亲和层析填料装入16/20层析柱;
(4)用20个柱体积的超纯水清洗柱子;
(5)清洗完成之后上10个柱体积的Buffer A冲洗;
(6)上清液与2×Buffer A按1:1的比例混合后调节pH至8.0,然后上镍亲和层析柱,流速5mL/min,上样后用Buffer A冲洗10个柱体积,再用层析缓冲液Buffer B洗脱,收集洗脱组分;
(7)收集得到的洗脱液使用30KDa 50mL的超滤浓缩管进行超滤,超滤至最后总体积为5mL,并将洗脱缓冲液置换成MOPS缓冲液,得到纯化后的蛋白。
图5示出了葡萄糖脱氢酶突变体Y406V与野生型FAD-GDH(WT)蛋白纯化的SDS-PAGE分析结果,其中七条泳道分别为:M:Marker;1:Y406V穿柱液;2:Y406V洗脱液;3:Y406V上清液;4:WT穿柱液;5:WT洗脱液;6:WT上清液。
按照实施例5中的方法对纯化后的蛋白进行酶活测定,发现纯化后,Y406V酶活达到8.8×10
实施例7.FAD-GDH突变体的酶学性质研究
1)底物专一性
进一步对实施例6精细纯化后得到的Y406V突变体和野生型FAD-GDH(WT)进行底物专一性比较,结果如表3所示。可以看出,Y406V对木糖和麦芽糖的底物专一性分别为:4.98±0.60%和1.31±0.40%。与野生型FAD-GDH相比P<0.05差异具有统计学意义。
表3.Y406V与FAD-GDH底物专一性比较
ND
2)最适反应pH值测定
将酶放入不同pH值的缓冲液中,测定酶活并进行比较,结果如图6所示,图6A为Y406V的酶活和pH值关系图,图6B为野生型FAD-GDH的酶活和pH值关系图。可以看出,毕赤酵母表达的Y406V在pH值6到8之间酶活较好,最适pH值为7.5,野生型FAD-GDH在pH值从7到8之间酶活力较好,最适pH值为8。
3)最适反应温度确定
在不同温度条件下测定酶活并比较,结果如图7所示,图7A为Y406V的酶活和温度关系图,图7B为野生型FAD-GDH的酶活和温度关系图。可以看出,突变体Y406V随着温度的升高酶活呈现下降的趋势,说明突变后的酶对温度的耐受减弱。野生型FAD-GDH的酶活在反应温度较低时随着反应温度的升高而提高,最适反应温度为55℃,温度继续升高酶活降低。
4)pH值稳定性
将Y406V与野生型FAD-GDH分别置于不同pH值的缓冲液中,在室温下放置1h,然后测定其残余的酶活,以此考察不同pH值对酶活的影响。结果如图8所示,图8A为Y406V的pH值稳定性图,图8B为野生型FAD-GDH的pH值稳定性图。可以看出,毕赤酵母表达的野生型FAD-GDH和Y406V具有相似的稳定pH值范围,而且突变体的范围更宽,Y406V大约在2.5到10之间,野生型FAD-GDH在3到10之间。
5)热稳定性
将Y406V与野生型FAD-GDH置于62℃条件下,检测不同时间下残余的酶活,结果如图9所示,图9A为Y406V的热稳定性图,图9B为野生型FAD-GDH的热稳定性图。可以看出,毕赤酵母表达的Y406V和野生型FAD-GDH具有较好的热稳定性,Y406V酶活的半衰期大约为25min,野生型FAD-GDH的热稳定性半衰期大约为35min。
6)酶促反应动力学性质
使用不同浓度的底物进行酶活测定,采用软件Graph-Pad Prism 7计算,可以得到相应的Km和Vmax值。对于毕赤酵母表达的Y406V,Km值为107±5.96mM高于野生型FAD-GDH的Km值99.95±4.83mM,其最大反应速率Vmax为1281±38U/mg,低于野生型FAD-GDH的最大反应速率Vmax值1414±24U/mg。
本领域技术人员应该理解的是,本发明的使用不受限于上述特定应用。就本文描述或描绘的特定元素和/或特征而言,本发明也不局限于其优选实施方案。应当理解的是,本发明不限于所公开的实施方案例或各个实施方案,且在不脱离由以下权利要求所阐述和限定的本发明的范围的情况下能够进行许多重新布置、修改和替换。
SEQUENCE LISTING
<110> 遵义医科大学珠海校区
<120> 一种葡萄糖脱氢酶突变体及其制备方法
<130> 111
<160> 6
<170> PatentIn version 3.5
<210> 1
<211> 574
<212> PRT
<213> Aspergillus terreus
<400> 1
Met Ser Asn Ser Thr Ser Ala Lys Tyr Asp Tyr Ile Val Ile Gly Gly
1 5 1015
Gly Thr Ser Gly Leu Ala Val Ala Asn Arg Leu Ser Glu Asp Pro Ser
202530
Val Asn Val Leu Ile Leu Glu Ala Gly Gly Ser Val Trp Asn Asn Pro
354045
Asn Val Thr Asn Val Asn Gly Tyr Gly Leu Ala Phe Gly Ser Asp Ile
505560
Asp Trp Gln Tyr Gln Ser Val Asn Gln Pro Tyr Gly Gly Asn Val Ser
65707580
Gln Val Leu Arg Ala Gly Lys Ala Leu Gly Gly Thr Ser Thr Ile Asn
859095
Gly Met Ala Tyr Thr Arg Ala Glu Asp Val Gln Ile Asp Ala Trp Glu
100 105 110
Thr Ile Gly Asn Thr Gly Trp Thr Trp Lys Asn Leu Phe Pro Tyr Tyr
115 120 125
Arg Lys Ser Glu Asn Phe Thr Val Pro Thr Lys Ser Gln Thr Ser Leu
130 135 140
Gly Ala Ser Tyr Glu Ala Gly Ala His Gly His Glu Gly Pro Leu Asp
145 150 155 160
Val Ala Phe Thr Gln Ile Glu Ser Asn Asn Leu Thr Thr Tyr Leu Asn
165 170 175
Arg Thr Phe Gln Gly Met Gly Leu Pro Trp Thr Glu Asp Val Asn Gly
180 185 190
Gly Lys Met Arg Gly Phe Asn Leu Tyr Pro Ser Thr Val Asn Leu Glu
195 200 205
Glu Tyr Val Arg Glu Asp Ala Ala Arg Ala Tyr Tyr Trp Pro Tyr Lys
210 215 220
Ser Arg Pro Asn Leu His Val Leu Leu Asn Thr Phe Ala Asn Arg Ile
225 230 235 240
Val Trp Asp Gly Glu Ala Arg Asp Gly Asp Ile Thr Ala Ser Gly Val
245 250 255
Glu Ile Thr Ser Arg Asn Gly Thr Val Arg Val Ile Asn Ala Glu Lys
260 265 270
Glu Val Ile Val Ser Ala Gly Ala Leu Lys Ser Pro Ala Ile Leu Glu
275 280 285
Leu Ser Gly Ile Gly Asn Pro Ser Val Leu Asp Lys Tyr Asn Ile Pro
290 295 300
Val Lys Val Asn Leu Pro Thr Val Gly Glu Asn Leu Gln Asp Gln Val
305 310 315 320
Asn Ser His Met Asp Ala Ser Gly Asn Thr Ser Ile Ser Gly Thr Lys
325 330 335
Ala Val Ser Tyr Pro Asp Val Tyr Asp Val Phe Gly Asp Glu Ala Glu
340 345 350
Ser Val Ala Lys Gln Ile Arg Ala Asn Leu Lys Gln Tyr Ala Ala Asp
355 360 365
Thr Ala Lys Ala Asn Gly Asn Ile Met Lys Ala Ala Asp Leu Glu Arg
370 375 380
Leu Phe Glu Val Gln Tyr Asp Leu Ile Phe Lys Gly Arg Val Pro Ile
385 390 395 400
Ala Glu Val Leu Asn Tyr Pro Gly Ser Ala Thr Ser Val Phe Ala Glu
405 410 415
Phe Trp Ala Leu Leu Pro Phe Ala Arg Gly Ser Val His Ile Gly Ser
420 425 430
Ser Asn Pro Ala Glu Phe Pro Val Ile Asn Pro Asn Tyr Phe Met Leu
435 440 445
Asp Trp Asp Ala Lys Ser Tyr Val Ala Val Ala Lys Tyr Ile Arg Arg
450 455 460
Ser Phe Glu Ser Tyr Pro Leu Ser Ser Ile Val Lys Glu Ser Thr Pro
465 470 475 480
Gly Tyr Asp Val Ile Pro Arg Asn Ala Ser Glu Gln Ser Trp Lys Glu
485 490 495
Trp Val Phe Asp Lys Asn Tyr Arg Ser Asn Phe His Pro Val Gly Thr
500 505 510
Ala Ala Met Met Pro Arg Glu Ile Gly Gly Val Val Asp Glu Arg Leu
515 520 525
Asn Val Tyr Gly Thr Thr Asn Val Arg Val Val Asp Ala Ser Val Leu
530 535 540
Pro Phe Gln Val Cys Gly His Leu Val Ser Thr Leu Tyr Ala Val Ala
545 550 555 560
Glu Arg Ala Ala Asp Leu Ile Lys Ala Asp Ala Gly Arg Arg
565 570
<210> 2
<211> 1725
<212> DNA
<213> Aspergillus terreus
<400> 2
atgtctaatt ctacgtctgc caagtacgat tacatcgtta ttggaggtgg cacatccggt 60
ttggctgttg ccaacagatt gtcagaggac ccttctgtaa atgtattaat cctggaagca 120
ggaggtagcg tatggaataa tcctaatgtg acgaacgtta acggatatgg actggcattt 180
ggatctgata ttgattggca atatcagtca gttaaccaac catacggtgg caacgtttca 240
caggtgttac gtgccggaaa agcattaggt ggcacatcca ccattaacgg aatggcatat 300
accagagctg aagatgtaca gattgatgcc tgggagacga ttggcaacac tggctggacc 360
tggaagaacc tgttcccata ttatcgtaag tcagaaaact ttacagtacc tacgaaatca 420
cagacaagcc tgggtgcatc atacgaggcc ggagcacatg gacatgaagg tcctctggat 480
gtggctttta cccagatcga atctaataat ttgactactt acttaaatag gacgtttcag 540
ggtatgggat taccttggac cgaagacgtt aacggtggaa aaatgagagg ttttaatctg 600
tatccatcaa ccgtgaacct ggaagagtat gtaagggagg atgccgccag agcctattat 660
tggccatata aatcaagacc aaatctgcat gtcttgctga atacattcgc caatcgtatt 720
gtatgggacg gagaggctcg tgatggtgat attaccgcaa gcggtgtcga aattacttcc 780
cgtaacggaa ccgttcgtgt tattaatgcc gaaaaagaag tcattgtcag cgccggcgcc 840
ctgaaatctc cagccatttt agagctgtct ggtatcggaa accctagcgt gttagacaag 900
tataacattc cagttaaggt aaacctgcct acggtgggtg aaaacctgca agatcaagtc 960
aactctcata tggatgcctc tggcaatact tcaatttcag gaaccaaagc agtgtcctac 1020
cctgacgttt atgatgtctt tggtgacgaa gctgaatctg ttgccaaaca gattcgtgct 1080
aatctgaagc aatatgctgc cgataccgca aaagccaatg gtaacatcat gaaggccgca 1140
gatctggagc gtctgttcga ggtccagtac gatttgatct tcaaaggtcg tgttccaatt 1200
gccgaagttt taaactatcc aggctccgcc accagcgtct tcgcagagtt ttgggctttg 1260
ttaccttttg cacgtggtag cgtccacatt ggatcttcaa atccagccga gtttccagtt 1320
attaacccaa actattttat gctggattgg gacgctaagt cttatgtggc agtcgccaag 1380
tatatcaggc gttctttcga aagctaccct ctgtcttcta tcgttaaaga aagcacccca 1440
ggatatgacg ttatccctag gaatgcatcc gagcagtcat ggaaagagtg ggtcttcgat 1500
aaaaattacc gttctaactt tcatcctgtt ggtactgccg caatgatgcc acgtgagatc 1560
ggcggtgttg tagacgagag actgaatgtt tacggtacga ccaacgtgag ggtagtagac 1620
gctagcgttt tgccattcca ggtgtgtggc cacctggtgt ctactctgta cgccgttgca 1680
gagagagccg ccgacttaat caaagccgat gctggcagac gttaa 1725
<210> 3
<211> 47
<212> DNA
<213> 人工序列(Artificial Sequence)
<400> 3
ctctcgagaa aagagaggct gaagcttcac acaggaaaca gaccatg 47
<210> 4
<211> 47
<212> DNA
<213> 人工序列(Artificial Sequence)
<400> 4
ggaacagtca tgtctaaggc gaattacatc cgccaaaaca gccaagc 47
<210> 5
<211> 33
<212> DNA
<213> 人工序列(Artificial Sequence)
<400> 5
tgccgaagtt ttaaacnnnc caggctccgc cac 33
<210> 6
<211> 33
<212> DNA
<213> 人工序列(Artificial Sequence)
<400> 6
gtggcggagc ctggnnngtt taaaacttcg gca 33
一种葡萄糖脱氢酶突变体及其制备方法专利购买费用说明
![]()
Q:办理专利转让的流程及所需资料
A:专利权人变更需要办理著录项目变更手续,有代理机构的,变更手续应当由代理机构办理。
1:专利变更应当使用专利局统一制作的“著录项目变更申报书”提出。
2:按规定缴纳著录项目变更手续费。
3:同时提交相关证明文件原件。
4:专利权转移的,变更后的专利权人委托新专利代理机构的,应当提交变更后的全体专利申请人签字或者盖章的委托书。
Q:专利著录项目变更费用如何缴交
A:(1)直接到国家知识产权局受理大厅收费窗口缴纳,(2)通过代办处缴纳,(3)通过邮局或者银行汇款,更多缴纳方式
Q:专利转让变更,多久能出结果
A:著录项目变更请求书递交后,一般1-2个月左右就会收到通知,国家知识产权局会下达《转让手续合格通知书》。















动态评分
0.0