

IPC分类号 : C12N1/21I,C12N15/70I,C12N15/53I,C12N15/54I,C12N15/61I,C12P23/00I,C12R1/19N



专利摘要
本发明公开一种基于杜氏藻代谢途径的β‑胡萝卜素高产工程菌及其构建方法与应用,涉及基因工程菌领域。本发明第一次实现全部利用来自杜氏藻(如杜氏巴氏藻或杜氏盐藻)的类胡萝卜素途径的基因来构建出高产β‑胡萝卜素的工程菌。当杜氏藻为杜氏巴氏藻时,所述β‑胡萝卜素高产工程菌的β‑胡萝卜素的产率为2.7mg/g(细胞干重);当杜氏藻为杜氏盐藻时,所述β‑胡萝卜素高产工程菌的β‑胡萝卜素的产率为3.3mg/g(细胞干重)。本发明采用的载体的启动子都是T7启动子,包含lacZ调控元件,其在IPTG作用的大肠杆菌BL21(DE3)中能够高表达,这有利于β‑胡萝卜素的大量积累,可用于实际应用生产。
权利要求
1.一种基于杜氏盐藻代谢途径的β-胡萝卜素高产工程菌,其特征在于:
所述β-胡萝卜素高产工程菌含有编码SEQ ID NO:10所示氨基酸序列的牦牛儿基牻牛儿基焦磷酸合成酶Ggps基因、编码GenBank:AAB51287.1所示氨基酸序列的八氢番茄红素合成酶Psy基因、编码GenBank:CAA75094.1所示氨基酸序列的八氢番茄红素脱氢酶Pds基因、编码SEQ ID NO:12所示氨基酸序列的15-顺式-ζ-胡萝卜素异构酶Ziso基因、编码SEQ IDNO:14所示氨基酸序列的ζ-胡萝卜素脱氢酶Zds基因、编码SEQ ID NO:16所示氨基酸序列的类胡萝卜素异构酶Crtiso基因和编码GenBank:ADX41685.1所示氨基酸序列的番茄红素β-环化酶Lycb基因。
2.根据权利要求1所述的基于杜氏盐藻代谢途径的β-胡萝卜素高产工程菌,其特征在于:
编码SEQ ID NO:10所示氨基酸序列的牦牛儿基牻牛儿基焦磷酸合成酶Ggps基因的核苷酸序列为如SEQ ID NO:9所示的核苷酸序列;
编码GenBank:AAB51287.1所示氨基酸序列的八氢番茄红素合成酶Psy基因的核苷酸序列为如GenBank:U91900.1所示的核苷酸序列;
编码GenBank:CAA75094.1所示氨基酸序列的八氢番茄红素脱氢酶Pds基因的核苷酸序列为如GenBank:Y14807.1所示的核苷酸序列;
编码SEQ ID NO:12所示氨基酸序列的15-顺式-ζ-胡萝卜素异构酶Ziso基因的核苷酸序列为如SEQ ID NO:11所示的核苷酸序列;
编码SEQ ID NO:14所示氨基酸序列的ζ-胡萝卜素脱氢酶Zds基因的核苷酸序列为如SEQ ID NO:13所示的核苷酸序列;
编码SEQ ID NO:16所示氨基酸序列的类胡萝卜素异构酶Crtiso基因的核苷酸序列为如SEQ ID NO:15所示的核苷酸序列;
编码GenBank:ADX41685.1所示氨基酸序列的番茄红素β-环化酶Lycb基因的核苷酸序列为如GenBank:HQ728089.1所示的核苷酸序列。
3.权利要求1~2任一项所述的基于杜氏盐藻代谢途径的β-胡萝卜素高产工程菌的构建方法,其特征在于,包括如下步骤:
(1)利用相关基因工程手段从杜氏盐藻中克隆到牦牛儿基牻牛儿基焦磷酸合成酶Ggps、八氢番茄红素合成酶Psy、八氢番茄红素脱氢酶Pds、15-顺式-ζ-胡萝卜素异构酶Ziso、ζ-胡萝卜素脱氢酶Zds、类胡萝卜素异构酶Crtiso、番茄红素β-环化酶Lycb;
(2)将Ggps和Psy构建在pACYduet-1载体上,氯霉素抗性,得到重组载体pACYduet-ggps-psy;将Pds、Zds和Lycb构建在pCDFduet-1载体上,链霉素抗性,得到重组载体pCDFduet-pds-zds-lycb;将Ziso和Crtiso构建在pETduet-1载体上,氨苄抗性,得到重组载体pETduet-ziso-crtiso;
(3)然后将步骤(2)构建的三个重组载体共转化到大肠杆菌BL21(DE3)中,获得基于杜氏盐藻代谢途径的β-胡萝卜素高产工程菌。
4.根据权利要求3所述的基于杜氏盐藻代谢途径的β-胡萝卜素高产工程菌的构建方法,其特征在于:
重组载体构建时,在7个目的基因的5′端均连接有核糖体结合位点rbs序列,或者,均连接有T7_promoter序列和核糖体结合位点rbs序列。
5.权利要求1~2任一项所述的基于杜氏盐藻代谢途径的β-胡萝卜素高产工程菌在生产β-胡萝卜素中的应用。
6.根据权利要求5所述的应用,特征在于:
所述β-胡萝卜素高产工程菌的β-胡萝卜素的产率为3.3 mg/g细胞干重。
说明书
技术领域
本发明涉及基因工程菌领域,涉及植物类胡萝卜素代谢途径载体的构建克隆系统,特别涉及一种基于杜氏藻代谢途径的β-胡萝卜素高产工程菌及其构建方法与应用;具体涉及一种杜氏藻(如杜氏巴氏藻(Dunaliella bardawil)、杜氏盐藻(Dunaliellasaline))中牦牛儿基牻牛儿基焦磷酸合成酶(Ggps)、八氢番茄红素合成酶(Psy)、八氢番茄红素脱氢酶(Pds)、15-顺式-ζ-胡萝卜素异构酶(Ziso)、ζ-胡萝卜素脱氢酶(Zds)、类胡萝卜素异构酶(Crtiso)、番茄红素β-环化酶(Lycb)基因的cDNA并构建出高产β-胡萝卜素的工程菌。
背景技术
类胡萝卜素分子是含8个异戊二烯单位的萜类化合物,其作为有机色素广泛存在于植物的叶绿体和有色体中,以及其它的一些光合组织中,比如藻类,部分细菌和真菌中。在特定条件下,杜氏藻细胞中类胡萝卜素含量可达到细胞干重的14%。其高含量可能跟酶的转化效率有很大的关系。
发明内容
为了克服现有技术的缺点与不足,本发明的首要目的在于提供一种基于杜氏藻代谢途径的β-胡萝卜素高产工程菌。
本发明的另一目的在于提供上述基于杜氏藻代谢途径的β-胡萝卜素高产工程菌的构建方法。
本发明的再一目的在于提供上述基于杜氏藻代谢途径的β-胡萝卜素高产工程菌的应用。
本发明的目的通过下述技术方案实现:
一种基于杜氏藻代谢途径的β-胡萝卜素高产工程菌,基于杜氏巴氏藻(Dunaliella bardawil)代谢途径时,所述β-胡萝卜素高产工程菌含有编码GenBank:APW83740.1所示氨基酸序列的牦牛儿基牻牛儿基焦磷酸合成酶(Ggps)基因、编码SEQ IDNO:2所示氨基酸序列的八氢番茄红素合成酶(Psy)基因、编码GenBank:ADD52599.1所示氨基酸序列的八氢番茄红素脱氢酶(Pds)基因、编码SEQ ID NO:4所示氨基酸序列的15-顺式-ζ-胡萝卜素异构酶(Ziso)基因、编码SEQ ID NO:6所示氨基酸序列的ζ-胡萝卜素脱氢酶(Zds)基因、编码SEQ ID NO:8所示氨基酸序列的类胡萝卜素异构酶(Crtiso)基因和编码GenBank:ANY98896.1所示氨基酸序列的番茄红素β-环化酶(Lycb)基因;
基于杜氏盐藻(Dunaliella saline)代谢途径时,所述β-胡萝卜素高产工程菌含有编码SEQ ID NO:10所示氨基酸序列的牦牛儿基牻牛儿基焦磷酸合成酶(Ggps)基因、编码GenBank:AAB51287.1所示氨基酸序列的八氢番茄红素合成酶(Psy)基因、编码GenBank:CAA75094.1所示氨基酸序列的八氢番茄红素脱氢酶(Pds)基因、编码SEQ ID NO:12所示氨基酸序列的15-顺式-ζ-胡萝卜素异构酶(Ziso)基因、编码SEQ ID NO:14所示氨基酸序列的ζ-胡萝卜素脱氢酶(Zds)基因、编码SEQ ID NO:16所示氨基酸序列的类胡萝卜素异构酶(Crtiso)基因和编码GenBank:ADX41685.1所示氨基酸序列的番茄红素β-环化酶(Lycb)基因;
优选的,基于杜氏巴氏藻(Dunaliella bardawil)代谢途径时,编码GenBank:APW83740.1所示氨基酸序列的牦牛儿基牻牛儿基焦磷酸合成酶(Ggps)基因的核苷酸序列为如GenBank:KX231795.1所示的核苷酸序列;其中,GenBank:APW83740.1与GenBank:KX231795.1中的来源Dunaliella salina记载有误,实际来源应该是Dunaliellabardawil;
编码SEQ ID NO:2所示氨基酸序列的八氢番茄红素合成酶(Psy)基因的核苷酸序列为如SEQ ID NO:1所示的核苷酸序列;
编码GenBank:ADD52599.1所示氨基酸序列的八氢番茄红素脱氢酶(Pds)基因的核苷酸序列为如GenBank:GQ923693.1所示的核苷酸序列;其中,GenBank:ADD52599.1与GenBank:GQ923693.1中的来源Dunaliella salina记载有误,实际来源应该是Dunaliellabardawil;
编码SEQ ID NO:4所示氨基酸序列的15-顺式-ζ-胡萝卜素异构酶(Ziso)基因的核苷酸序列为如SEQ ID NO:3所示的核苷酸序列;
编码SEQ ID NO:6所示氨基酸序列的ζ-胡萝卜素脱氢酶(Zds)基因的核苷酸序列为如SEQ ID NO:5所示的核苷酸序列;
编码SEQ ID NO:8所示氨基酸序列的类胡萝卜素异构酶(Crtiso)基因的核苷酸序列为如SEQ ID NO:7所示的核苷酸序列;
编码GenBank:ANY98896.1所示氨基酸序列的番茄红素β-环化酶(Lycb)基因的核苷酸序列为如GenBank:KX218392.1所示的核苷酸序列;其中,GenBank:ANY98896.1与GenBank:KX218392.1中的来源Dunaliella salina记载有误,实际来源应该是Dunaliellabardawil;
优选的,基于杜氏盐藻(Dunaliella saline)代谢途径时,编码SEQ ID NO:10所示氨基酸序列的牦牛儿基牻牛儿基焦磷酸合成酶(Ggps)基因的核苷酸序列为如SEQ ID NO:9所示的核苷酸序列;
编码GenBank:AAB51287.1所示氨基酸序列的八氢番茄红素合成酶(Psy)基因的核苷酸序列为如GenBank:U91900.1所示的核苷酸序列;其中,GenBank:U91900.1中的来源Dunaliella bardawil记载有误,实际来源应该是Dunaliella saline;
编码GenBank:CAA75094.1所示氨基酸序列的八氢番茄红素脱氢酶(Pds)基因的核苷酸序列为如GenBank:Y14807.1所示的核苷酸序列;其中,GenBank:Y14807.1中的来源Dunaliella bardawil记载有误,实际来源应该是Dunaliella saline;
编码SEQ ID NO:12所示氨基酸序列的15-顺式-ζ-胡萝卜素异构酶(Ziso)基因的核苷酸序列为如SEQ ID NO:11所示的核苷酸序列;
编码SEQ ID NO:14所示氨基酸序列的ζ-胡萝卜素脱氢酶(Zds)基因的核苷酸序列为如SEQ ID NO:13所示的核苷酸序列;
编码SEQ ID NO:16所示氨基酸序列的类胡萝卜素异构酶(Crtiso)基因的核苷酸序列为如SEQ ID NO:15所示的核苷酸序列;
编码GenBank:ADX41685.1所示氨基酸序列的番茄红素β-环化酶(Lycb)基因的核苷酸序列为如GenBank:HQ728089.1所示的核苷酸序列。
所述的基于杜氏藻代谢途径的β-胡萝卜素高产工程菌的构建方法,包括如下步骤:
(1)利用相关基因工程手段从杜氏藻中克隆到牦牛儿基牻牛儿基焦磷酸合成酶(Ggps)、八氢番茄红素合成酶(Psy)、八氢番茄红素脱氢酶(Pds)、15-顺式-ζ-胡萝卜素异构酶(Ziso)、ζ-胡萝卜素脱氢酶(Zds)、类胡萝卜素异构酶(Crtiso)、番茄红素β-环化酶(Lycb);
(2)将Ggps和Psy构建在pACYduet-1载体上,氯霉素抗性,得到重组载体pACYduet-ggps-psy;将Pds、Zds和Lycb构建在pCDFduet-1载体上,链霉素抗性,得到重组载体pCDFduet-pds-zds-lycb;将Ziso和Crtiso构建在pETduet-1载体上,氨苄抗性,得到重组载体pETduet-ziso-crtiso;
(3)然后将步骤(2)构建的三个重组载体共转化到大肠杆菌BL21(DE3)中,获得基于杜氏藻代谢途径的β-胡萝卜素高产工程菌。
优选的,所述的杜氏藻为杜氏巴氏藻(Dunaliella bardawil)或杜氏盐藻(Dunaliella saline)。
优选的,重组载体构建时,在7个目的基因的N端(5′端)均连接有rbs(核糖体结合位点)序列,或者,均连接有T7启动子(T7_promoter)序列和rbs(核糖体结合位点)序列。
所述的基于杜氏藻代谢途径的β-胡萝卜素高产工程菌在生产β-胡萝卜素中的应用。
当杜氏藻为杜氏巴氏藻(Dunaliella bardawil)时,所述β-胡萝卜素高产工程菌的β-胡萝卜素的产率为2.7mg/g(细胞干重)。
当杜氏藻为杜氏盐藻(Dunaliella saline)时,所述β-胡萝卜素高产工程菌的β-胡萝卜素的产率为3.3mg/g(细胞干重)。
本发明相对于现有技术具有如下的优点及效果:
本发明第一次实现全部利用来自杜氏藻(如杜氏巴氏藻或杜氏盐藻)的类胡萝卜素途径的基因来构建工程菌。而杜氏藻(如杜氏巴氏藻或杜氏盐藻)是已知的产β-胡萝卜素最高的植物之一。本发明采用的载体的启动子都是T7启动子,包含lacZ调控元件,其在IPTG作用的大肠杆菌BL21(DE3)中能够高表达,这有利于β-胡萝卜素的大量积累,可用于实际应用生产。
附图说明
图1是本发明pACYduet-Dbggps-Dbpsy(A)和pCDFduet-Dbpds-Dbzds-Dblycb(B)构建图。
图2是本发明pETduet-Dbziso-Dbcrtiso构建图。
图3是本发明基于杜氏藻代谢途径的β-胡萝卜素高产工程菌与普通大肠杆菌BL21(DE3)颜色比较;其中,A是基于杜氏巴氏藻代谢途径的β-胡萝卜素高产工程菌;B是基于杜氏盐藻代谢途径的β-胡萝卜素高产工程菌。
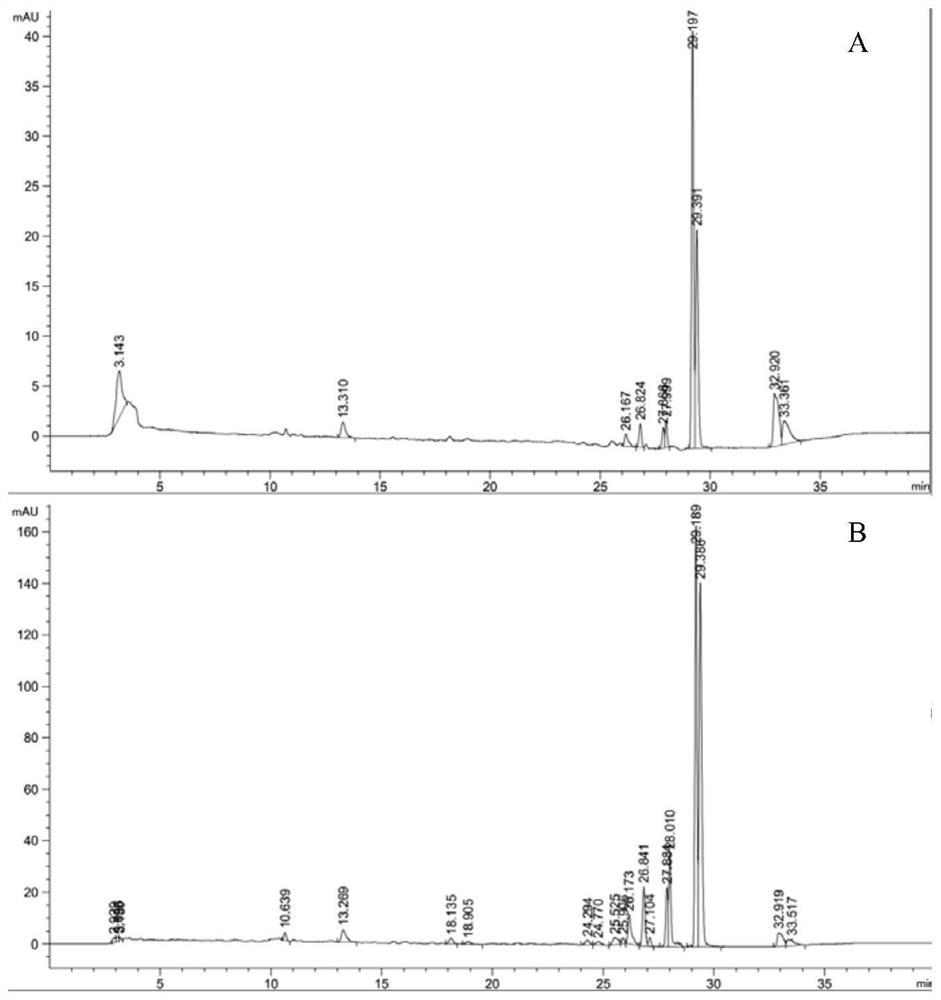
图4是β-胡萝卜素液相图;其中,A是实施例1;B是实施例2。
图5是本发明pACYduet1-Dsggps-Dspsy构建图。
图6是本发明pCDFduet1-Dspds-Dszds-Dslycb构建图。
图7是本发明pETduet1-Dsziso-Dscrtiso构建图。
具体实施方式
下面结合实施例及附图对本发明作进一步详细的描述,但本发明的实施方式不限于此。
下列实施例中未注明具体实验条件的试验方法,通常按照常规实验条件或按照制造厂所建议的实验条件。所使用的材料、试剂等,如无特殊说明,为从商业途径得到的试剂和材料。
实施例1
1.制备线性化载体
(1)用NcoI和HindIII(购自Thermo Fisher Scientific)双酶切质粒pACYduet-1,pCDFduet-1和pETduet-1(购自EMD Biosciences)。37℃水浴3h。酶切反应体系如下:
10×Buffer 5μL、两种酶各1μL、质粒10μL、Add dH2O To 50μL。
(2)1%琼脂糖凝胶电泳检测酶切产物。
(3)酶切之后,采用PCR纯化试剂盒纯化线性化载体。
2.PCR扩增目的片段
(1)以杜氏巴氏藻(Dunaliella bardawil)(购自中科院水生生物研究所淡水藻种库,编号FACHB-847)反转录的cDNA为模板,使用Takara的PrimeSTAR HS DNA Polymerase(高保真酶,购自Takara),进行PCR反应。
(2)1%琼脂糖凝胶电泳检测PCR产物。
(3)用OMEGA公司的胶回收试剂盒E.Z.N.A.TM Gel Extraction Kit回收目的片段,纯化PCR产物。测序结果如GenBank:KX231795.1、SEQ ID NO:1、GenBank:GQ923693.1、SEQ ID NO:3、5、7、GenBank:KX218392.1所示,对应编码的氨基酸序列如GenBank:APW83740.1、SEQ ID NO:2、GenBank:ADD52599.1、SEQ ID NO:4、6、8、GenBank:ANY98896.1所示。
3.In-Fusion克隆
用NcoI和HindIII双酶切质粒pACYduet-1并插入用引物Pac_dbggps_F和Pac_dbggps_R扩增得到的Dbggps,得到pACYduet-Dbggps;再用NdeI和EcoRV双酶切pACYduet-Dbggps并插入用引物Pac_dbpsy_F和Pac_dbpsy_R扩增得到的Dbpsy,得到pACYduet-Dbggps-Dbpsy。引物序列如表1所示。
用NcoI和HindIII双酶切质粒pCDFduet-1并插入用引物Pcd_dbpds_F和Pcd_dbpds_R扩增得到的Dbpds,得到pCDFduet-Dbpds;再用NdeI和EcoRV双酶切pCDFduet-Dbpds并插入用引物Pcd_dbzds_F和Pcd_dbzds_R扩增得到的Dbzds,得到pCDFduet-Dbpds-Dbzds;再用AatII和KpnI双酶切pCDFduet-Dbpds-Dbzds并插入用引物Pcd_dblycb_F和Pcd_dblycb_R扩增得到的Dblycb,得到pCDFduet-Dbpds-Dbzds-Dblycb。引物序列如表1所示。
用NcoI和HindIII双酶切质粒pETduet-1并插入用引物Pet_dbziso_F和Pet_dbziso_R扩增得到的Dbziso,得到pETduet-Dbziso;再用NdeI和EcoRV双酶切pETduet-Dbziso并插入用引物Pet_dbcrtiso_F和Pet_dbcrtiso_R扩增得到的Dbcrtiso,得到pETduet-Dbziso-Dbcrtiso。引物序列如表1所示。
(1)建立In-Fusion(购自Takara)克隆反应体系:
(2)50℃孵育15min,然后置于冰上。
(3)连接产物分别转化大肠杆菌DH5α,涂布于含有氯霉素(CmR或CamR)、链霉素(Sm)或氨苄青霉素(Ap或Amp)的相应平板上。在37℃生化培养箱中分别倒置培养12~16h。注意培养时间不宜过长,否则抗性失效,生长出卫星菌落,给筛选阳性克隆造成困难。
表1构建重组质粒的引物
4.质粒的提取
参照OMEGA公司的E.Z.N.A.TM Plasmid Mini Kit试剂盒的说明书,提取重组质粒pACYduet-Dbggps-Dbpsy(图1A)、pCDFduet-Dbpds-Dbzds-Dblycb(图1B)、pETduet-Dbziso-Dbcrtiso(图2)。
(1)10000g,1min离心,收集菌体。(2)加入250μL SolutionⅠ,涡旋十几秒,混匀。(3)加入250μL SolutionⅡ,上下颠倒4~6次,室温放置2min。(4)加入350μL SolutionⅢ,轻轻摇匀,10000g,10min离心。(5)转上清至Hibind柱,10000g,1min离心。(6)弃掉滤液,加入500μL Buffer HB,10000g,1min离心。(7)弃掉滤液,加入700μL DNA Wash Buffer,10000g,1min离心。(8)重复洗一次(步骤(7))。(9)10000g,空管离心2min。(10)弃掉收集管将HiBind柱套进干净的1.5mL EP管,加入50~100μL灭菌水(60℃预热)洗脱DNA,静置1~2min后,10000g,1min离心。
5.转化大肠杆菌BL21(DE3)
将提取的重组质粒pACYduet-Dbggps-Dbpsy、pCDFduet-Dbpds-Dbzds-Dblycb和pETduet-Dbziso-Dbcrtiso共转化到大肠杆菌BL21(DE3),涂布于含有CmR、Sm和Ap抗生素的平板上,在37℃生化培养箱中倒置培养12~16h。经过筛选获得目的菌株,即基于杜氏巴氏藻代谢途径的β-胡萝卜素高产工程菌,如图3A所示,白色为BL21(DE3)对照菌株,土黄色的为高产β-胡萝卜素菌株。
6.转化子的培养
将目的菌株接种于50mL已灭菌的LB液体培养基中(加入氯霉素、氨苄青霉素、链霉素抗性),于200rpm,37℃下振荡培养6h。加入IPTG进行诱导,培养6小时。
7.提取转化子的类胡萝卜素
(1)将24mL菌液于4℃,8000rpm下离心2min,弃上清。(2)用去离子水重新悬浮大肠杆菌细胞,4℃,8000rpm离心2min,弃上清。(3)重复步骤(2)。(4)加6mL丙酮至大肠杆菌沉淀,涡旋振荡分散。(5)55℃水浴20min,每隔5min振荡一次。(6)将混合物于4℃,8000rpm下离心20min,收集上清。(7)用丙酮定容至6mL,-20℃保存。
8.HPLC定性分析转化子的类胡萝卜素成分
(1)将流动相用0.22μm孔径滤膜进行抽滤处理,接着超声处理60min去除液体中的气泡。
(2)将类胡萝卜素提取物以及β-胡萝卜素标准品溶液,用0.2μm孔径的聚碳酸酯滤膜进行过滤处理。
(3)调节色谱柱(C30YMC carotenoid column,5μm,250*4.6mm)温度在25℃,以1.0mL/min的流速用甲醇冲洗至平衡(约2h)。
(4)改为用流动相甲基叔丁基醚(A相)/甲醇(B相)(梯度洗脱:0min,90%A相和10%B相;10min,60%A相和40%B相;20min 50%A相和50%B相;25min,10%A相和90%B相;29.5min,90%A相和10%B相),以1mL/min的流速进行洗柱,直至平衡。
(5)将浓度为100μg/mL的β-胡萝卜素标准品存储液稀释为0.625μg/mL、1.25μg/mL、2.5μg/mL、5μg/mL、10μg/mL和20μg/mL等不同浓度的工作液,用HPLC仪测定473nm波长下工作液的峰面积,根据结果绘制标准曲线。
(6)上样20μL,在波长473nm下检测40min。
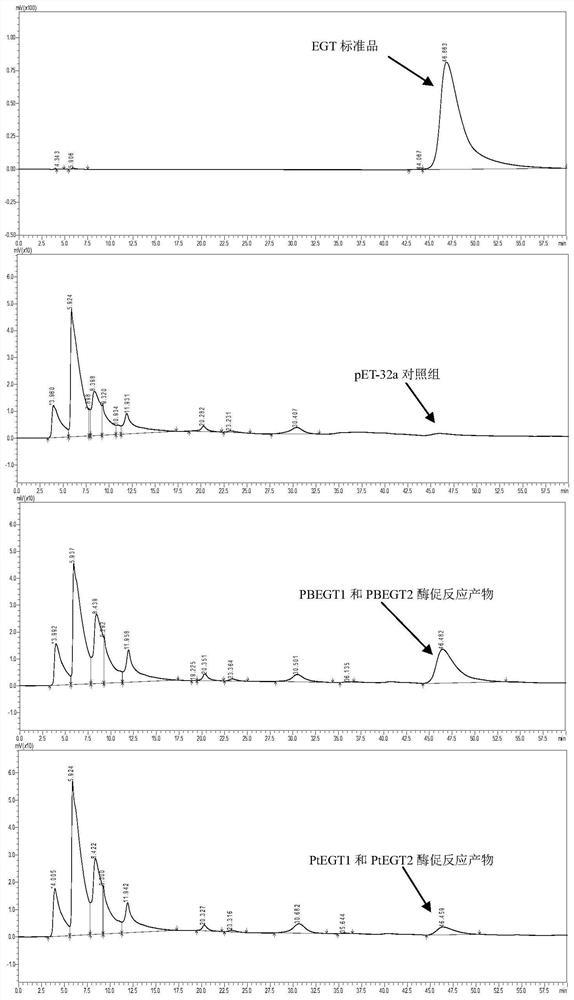
结果如图4A所示,表明在16.2min左右出现了β-胡萝卜素的特征峰,这与标准品的出峰时间是一致的,另外根据该峰面积,结合得到的标准曲线方程:y=34.791x+59.973(x为峰面积,y为β-胡萝卜素的含量),再根据细胞的干重,可以计算出β-胡萝卜素的产率为2.7mg/g(细胞干重)。
实施例2
1.制备线性化载体
(1)用Asc I和Sal I(购自Thermo Fisher Scientific)双酶切质粒pACYduet-1,pCDFduet-1和pETduet-1(购自EMD Biosciences)。37℃水浴3h。酶切反应体系如下:
10×Buffer 5μL、两种酶各1μL、质粒10μL、Add dH2O To 50μL。
(2)1%琼脂糖凝胶电泳检测酶切产物。
(3)酶切之后,采用PCR纯化试剂盒纯化线性化载体。
2.PCR扩增目的片段
(1)以杜氏盐藻(Dunaliella saline)(购自Culture Collection of Algae andProtozoa,英国;编号CCAP19/18)反转录的cDNA为模板,使用Takara的PrimeSTAR HS DNAPolymerase(高保真酶,购自Takara),进行PCR反应。
(2)1%琼脂糖凝胶电泳检测PCR产物。
(3)用OMEGA公司的胶回收试剂盒E.Z.N.A.TM Gel Extraction Kit回收目的片段,纯化PCR产物。测序结果如SEQ ID NO:9、GenBank:U91900.1、GenBank:Y14807.1、SEQ IDNO:11、13、15、GenBank:HQ728089.1所示,对应编码的氨基酸序列如SEQ ID NO:10、GenBank:AAB51287.1、GenBank:CAA75094.1、SEQ ID NO:12、14、16、GenBank:ADX41685.1所示。
3.In-Fusion克隆
用Asc I和Sal I双酶切质粒pACYduet-1并插入用引物Pac_dsggps_F和Pac_dsggps_R扩增得到的Dsggps,得到pACYduet1-Dsggps;再用NdeI和EcoRV双酶切pACYduet1-Dsggps并插入用引物Pac_dspsy_F和Pac_dspsy_R扩增得到的Dspsy,得到pACYduet1-Dsggps-Dspsy。引物序列如表2所示。
用Asc I和Sal I双酶切质粒pCDFduet-1并插入用引物Pcd_dspds_F和Pcd_dspds_R扩增得到的Dspds,得到pCDFduet1-Dspds;再用NdeI和EcoRV双酶切pCDFduet1-Dspds并插入用引物Pcd_dszds_F和Pcd_dszds_R扩增得到的Dszds,得到pCDFduet1-Dspds-Dszds;再用AatII和KpnI双酶切pCDFduet1-Dspds-Dszds并插入用引物Pcd_dslycb_F和Pcd_dslycb_R扩增得到的Dslycb,得到pCDFduet1-Dspds-Dszds-Dslycb。引物序列如表2所示。
用Asc I和Sal I双酶切质粒pETduet-1并插入用引物Pet_dsziso_F和Pet_dsziso_R扩增得到的Dsziso,得到pETduet1-Dsziso;再用NdeI和EcoRV双酶切pETduet1-Dsziso并插入用引物Pet_dscrtiso_F和Pet_dscrtiso_R扩增得到的Dscrtiso,得到pETduet1-Dsziso-Dscrtiso。引物序列如表2所示。
(1)建立In-Fusion(购自Takara)克隆反应体系:
(2)50℃孵育15min,然后置于冰上。
(3)连接产物分别转化大肠杆菌DH5α,涂布于含有氯霉素(CmR或CamR)、链霉素(Sm)或氨苄青霉素(Ap或Amp)的相应的平板上。在37℃生化培养箱中分别倒置培养12~16h。注意培养时间不宜过长,否则抗性失效,生长出卫星菌落,给筛选阳性克隆造成困难。
表2构建重组质粒的引物
4.质粒的提取
参照OMEGA公司的E.Z.N.A.TM Plasmid Mini Kit试剂盒的说明书,提取重组质粒pACYduet1-Dsggps-Dspsy(图5)、pCDFduet1-Dspds-Dszds-Dslycb(图6)、pETduet1-Dsziso-Dscrtiso(图7)。
(1)10000g,1min离心,收集菌体。(2)加入250μL SolutionⅠ,涡旋十几秒,混匀。(3)加入250μL SolutionⅡ,上下颠倒4~6次,室温放置2min。(4)加入350μL SolutionⅢ,轻轻摇匀,10000g,10min离心。(5)转上清至Hibind柱,10000g,1min离心。(6)弃掉滤液,加入500μL Buffer HB,10000g,1min离心。(7)弃掉滤液,加入700μL DNA Wash Buffer,10000g,1min离心。(8)重复洗一次(步骤(7))。(9)10000g,空管离心2min。(10)弃掉收集管将HiBind柱套进干净的1.5mL EP管,加入50~100μL灭菌水(60℃预热)洗脱DNA,静置1~2min后,10000g,1min离心。
5.转化大肠杆菌BL21(DE3)
将提取的重组质粒pACYduet1-Dsggps-Dspsy、pCDFduet1-Dspds-Dszds-Dslycb和pETduet1-Dsziso-Dscrtiso共转化到大肠杆菌BL21(DE3),涂布于含有CamR、Sm和Amp抗生素的平板上,在37℃生化培养箱中倒置培养12~16h。经过筛选获得目的菌株,即基于杜氏盐藻代谢途径的β-胡萝卜素高产工程菌,如图3B所示,白色为BL21(DE3)对照菌株,土黄色的为高产β-胡萝卜素菌株。
6.转化子的培养
将目的菌株接种于50mL已灭菌的LB液体培养基中(加入氯霉素、氨苄青霉素、链霉素抗性),于200rpm,37℃下振荡培养6h。加入IPTG进行诱导,培养6小时。
7.提取转化子的类胡萝卜素
(1)将24mL菌液于4℃,8000rpm下离心2min,弃上清。(2)用去离子水重新悬浮大肠杆菌细胞,4℃,8000rpm离心2min,弃上清。(3)重复步骤(2)。(4)加6mL丙酮至大肠杆菌沉淀,涡旋振荡分散。(5)55℃水浴20min,每隔5min振荡一次。(6)将混合物于4℃,8000rpm下离心20min,收集上清。(7)用丙酮定容至6mL,-20℃保存。
8.HPLC定性分析转化子的类胡萝卜素成分
(1)将流动相用0.22μm孔径滤膜进行抽滤处理,接着超声处理60min去除液体中的气泡。
(2)将类胡萝卜素提取物以及β-胡萝卜素标准品溶液,用0.2μm孔径的聚碳酸酯滤膜进行过滤处理。
(3)调节色谱柱(C30YMC carotenoid column,5μm,250*4.6mm)温度在25℃,以1.0mL/min的流速用甲醇冲洗至平衡(约2h)。
(4)改为用流动相甲基叔丁基醚(A相)/甲醇(B相)(梯度洗脱:0min,90%A相和10%B相;10min,60%A相和40%B相;20min 50%A相和50%B相;25min,10%A相和90%B相;29.5min,90%A相和10%B相),以1mL/min的流速进行洗柱,直至平衡。
(5)β-胡萝卜素的标准曲线参照实施例1步骤8(5)。
(6)上样20μL,在波长473nm下检测40min。
结果如图4B所示,表明在16.2min左右出现了β-胡萝卜素的特征峰,这与标准品的出峰时间是一致的,另外根据该峰面积,结合得到的标准曲线方程:y=34.791x+59.973(x为峰面积,y为β-胡萝卜素的含量),再根据细胞的干重,可以计算出β-胡萝卜素的产率为3.3mg/g(细胞干重)。
上述实施例为本发明较佳的实施方式,但本发明的实施方式并不受上述实施例的限制,其他的任何未背离本发明的精神实质与原理下所作的改变、修饰、替代、组合、简化,均应为等效的置换方式,都包含在本发明的保护范围之内。
序列表
<110> 华南理工大学
<120> 基于杜氏藻代谢途径的β-胡萝卜素高产工程菌及其构建方法与应用
<160> 16
<170> SIPOSequenceListing 1.0
<211> 1305
<212> DNA
<213> 人工序列(Artificial Sequence)
<220>
<223> 杜氏巴氏藻(Dunaliella bardawil):Psy
<400> 1
atgacgctgt caatgttgga cgcgcgaagg atggcacagc gaacagcaac ttcctcctcc 60
tcctctccta gcatcatata tgccccatcg cccataagca atcgcagcgg caggcgcgca 120
gcagcgaatc acggcatcag gaatggtagt cgcagagcag caggccggat gggactctgc 180
agcactgtgc aagtgaactg cacgctcgcg atgccccagc ccaaccacgg ccagaagatg 240
cgattgcagc agcagcagca gcaacaactg cagcagcagc agcaacagca actatcggga 300
aagcaggtgg aggagcaggc gatgctgcag tgcataaaga ccgctcagtc agtgcccccc 360
tccaccggac tcctcaatcc tcgcggcctg cgatggcagg gcagcagctt ggaagcagcg 420
tacgagcgat gtggggcggt gtgcagcgag tacgccaaga ccttctacct cggtactcag 480
ctcatgacac cagtgcaggc caggtgcatc tgggccatct atgtgtggtg ccgccgcaca 540
gatgagctgg tggatggccc caatgcatca aagatcacgc ctcaggccct agacagatgg 600
gaggagcgcc ttgaaagcat gttccaaggc aagccctatg atgtgctgga cgcggcgctc 660
acagacacca tctccaaatt ccctctggag gtgcaaccct tcagagacat gatcgagggc 720
atgcgaatgg acctcttcaa gtcgcggtat cacacctttg atgagctgta cgagtactgc 780
tatcgtgtgg cgggcacagt ggggctgatg accatgccag tgatggggat tgatcccaac 840
tacaagggtc caattgacaa ggtctacaag gccgcccttg cgctgggtac ggcaaaccag 900
ctcaccaaca ttctgcgaga tgtgggagag gacatcagag agcgtgaccg tatctacttg 960
cccctggatg agctcaagca gttcggcatc tccgaagagg aggtaaaagc aggtatccac 1020
aagccatcgc aaggcaaggt ggatgagcgg tggcgagcgt tcatgaagtt ccagatcaag 1080
cgtgcgcgag agtacttcca ggaagcagag gatggggtag actacttgga cgtgaaggcg 1140
cggtggccag tgtggtcagc gctgatcctt taccgccaaa tcttggatgt cattgagaag 1200
aatgactacg acaacttctc catgcgcgca tacgtgccta agtccaagaa gtttgcatcg 1260
ttgccgatgg ccttgttccg ggccttggtg cccaagaaca aataa 1305
<211> 434
<212> PRT
<213> 人工序列(Artificial Sequence)
<220>
<223> 杜氏巴氏藻(Dunaliella bardawil):Psy
<400> 2
Met Thr Leu Ser Met Leu Asp Ala Arg Arg Met Ala Gln Arg Thr Ala
1 5 10 15
Thr Ser Ser Ser Ser Ser Pro Ser Ile Ile Tyr Ala Pro Ser Pro Ile
20 25 30
Ser Asn Arg Ser Gly Arg Arg Ala Ala Ala Asn His Gly Ile Arg Asn
35 40 45
Gly Ser Arg Arg Ala Ala Gly Arg Met Gly Leu Cys Ser Thr Val Gln
50 55 60
Val Asn Cys Thr Leu Ala Met Pro Gln Pro Asn His Gly Gln Lys Met
65 70 75 80
Arg Leu Gln Gln Gln Gln Gln Gln Gln Leu Gln Gln Gln Gln Gln Gln
85 90 95
Gln Leu Ser Gly Lys Gln Val Glu Glu Gln Ala Met Leu Gln Cys Ile
100 105 110
Lys Thr Ala Gln Ser Val Pro Pro Ser Thr Gly Leu Leu Asn Pro Arg
115 120 125
Gly Leu Arg Trp Gln Gly Ser Ser Leu Glu Ala Ala Tyr Glu Arg Cys
130 135 140
Gly Ala Val Cys Ser Glu Tyr Ala Lys Thr Phe Tyr Leu Gly Thr Gln
145 150 155 160
Leu Met Thr Pro Val Gln Ala Arg Cys Ile Trp Ala Ile Tyr Val Trp
165 170 175
Cys Arg Arg Thr Asp Glu Leu Val Asp Gly Pro Asn Ala Ser Lys Ile
180 185 190
Thr Pro Gln Ala Leu Asp Arg Trp Glu Glu Arg Leu Glu Ser Met Phe
195 200 205
Gln Gly Lys Pro Tyr Asp Val Leu Asp Ala Ala Leu Thr Asp Thr Ile
210 215 220
Ser Lys Phe Pro Leu Glu Val Gln Pro Phe Arg Asp Met Ile Glu Gly
225 230 235 240
Met Arg Met Asp Leu Phe Lys Ser Arg Tyr His Thr Phe Asp Glu Leu
245 250 255
Tyr Glu Tyr Cys Tyr Arg Val Ala Gly Thr Val Gly Leu Met Thr Met
260 265 270
Pro Val Met Gly Ile Asp Pro Asn Tyr Lys Gly Pro Ile Asp Lys Val
275 280 285
Tyr Lys Ala Ala Leu Ala Leu Gly Thr Ala Asn Gln Leu Thr Asn Ile
290 295 300
Leu Arg Asp Val Gly Glu Asp Ile Arg Glu Arg Asp Arg Ile Tyr Leu
305 310 315 320
Pro Leu Asp Glu Leu Lys Gln Phe Gly Ile Ser Glu Glu Glu Val Lys
325 330 335
Ala Gly Ile His Lys Pro Ser Gln Gly Lys Val Asp Glu Arg Trp Arg
340 345 350
Ala Phe Met Lys Phe Gln Ile Lys Arg Ala Arg Glu Tyr Phe Gln Glu
355 360 365
Ala Glu Asp Gly Val Asp Tyr Leu Asp Val Lys Ala Arg Trp Pro Val
370 375 380
Trp Ser Ala Leu Ile Leu Tyr Arg Gln Ile Leu Asp Val Ile Glu Lys
385 390 395 400
Asn Asp Tyr Asp Asn Phe Ser Met Arg Ala Tyr Val Pro Lys Ser Lys
405 410 415
Lys Phe Ala Ser Leu Pro Met Ala Leu Phe Arg Ala Leu Val Pro Lys
420 425 430
Asn Lys
<211> 1104
<212> DNA
<213> 人工序列(Artificial Sequence)
<220>
<223> 杜氏巴氏藻(Dunaliella bardawil):Ziso
<400> 3
atggcgagct tgtgtagagc tgcccttggg caggccagtg cgaaggggct gagtggctta 60
caaacttcct ccaagcctct catctccaag agccctcttg tagcaagacc accatgcatc 120
agaattagcg aacgctgccc agtcttagaa aattccgtat ggagcagaag acgagcggag 180
gtgtgtgtgc gggctgctag cgacgaggag tcgccccgac ctgcggggct tgttggcgag 240
gatgcggcag ccttcgacgt ttcccagcag agtaccaagt cctgggcgct gtttactggg 300
cttctgactg gcgtgctggg cctcatttac ctggtatgga tccagccggg agcaggtctg 360
gcagatgact tcttggggtt ggtcgagggc ttcagcaaca acaacccgga ggcaacaatc 420
ctccttatcc tctttgtgtt tgctgttgta cacagtgggc tggcaggcct gcgccccaaa 480
ggggagcagc tgatcggcgc acgagcatac cgggtgattt ttgcccttgt cagcctgccc 540
ttggccatcg tggccatcgt gtacttcatc aaccatagat acgatggcat gcccttatgg 600
gatctcaggg gtgtgacggg agtgcatgag ctggtgtggc tcctcaactt tgtgtccttc 660
tacttccttt acccttccac ctttaacatc cttgaggtgg ctgcagtgga cgagcccaag 720
ctgcatatgt gggaaacggg aatcatgcga atcactcgtc acccacaaat ggtagggcag 780
ctaatatggt gcgcagcaca cacgttgtgg atcgggaaca gcttcatgct cgtgacctca 840
gcaggcctca tggctcatca tctctttggt tgctggcatg gtgaccggcg gttgtctgcc 900
aagtatggcg aggcctttga aatcgtcaag gcgcggacaa gtacctttcc attgcaagct 960
atttgggagg gccgccaggt cttgccagct gattactaca aggagttcct gcgggcgccc 1020
tacttcgctg tgactgcctt caccctgggc gcctacttcg cacaccccat catgcagtct 1080
gccagcttct atttgagatg gtag 1104
<211> 367
<212> PRT
<213> 人工序列(Artificial Sequence)
<220>
<223> 杜氏巴氏藻(Dunaliella bardawil):Ziso
<400> 4
Met Ala Ser Leu Cys Arg Ala Ala Leu Gly Gln Ala Ser Ala Lys Gly
1 5 10 15
Leu Ser Gly Leu Gln Thr Ser Ser Lys Pro Leu Ile Ser Lys Ser Pro
20 25 30
Leu Val Ala Arg Pro Pro Cys Ile Arg Ile Ser Glu Arg Cys Pro Val
35 40 45
Leu Glu Asn Ser Val Trp Ser Arg Arg Arg Ala Glu Val Cys Val Arg
50 55 60
Ala Ala Ser Asp Glu Glu Ser Pro Arg Pro Ala Gly Leu Val Gly Glu
65 70 75 80
Asp Ala Ala Ala Phe Asp Val Ser Gln Gln Ser Thr Lys Ser Trp Ala
85 90 95
Leu Phe Thr Gly Leu Leu Thr Gly Val Leu Gly Leu Ile Tyr Leu Val
100 105 110
Trp Ile Gln Pro Gly Ala Gly Leu Ala Asp Asp Phe Leu Gly Leu Val
115 120 125
Glu Gly Phe Ser Asn Asn Asn Pro Glu Ala Thr Ile Leu Leu Ile Leu
130 135 140
Phe Val Phe Ala Val Val His Ser Gly Leu Ala Gly Leu Arg Pro Lys
145 150 155 160
Gly Glu Gln Leu Ile Gly Ala Arg Ala Tyr Arg Val Ile Phe Ala Leu
165 170 175
Val Ser Leu Pro Leu Ala Ile Val Ala Ile Val Tyr Phe Ile Asn His
180 185 190
Arg Tyr Asp Gly Met Pro Leu Trp Asp Leu Arg Gly Val Thr Gly Val
195 200 205
His Glu Leu Val Trp Leu Leu Asn Phe Val Ser Phe Tyr Phe Leu Tyr
210 215 220
Pro Ser Thr Phe Asn Ile Leu Glu Val Ala Ala Val Asp Glu Pro Lys
225 230 235 240
Leu His Met Trp Glu Thr Gly Ile Met Arg Ile Thr Arg His Pro Gln
245 250 255
Met Val Gly Gln Leu Ile Trp Cys Ala Ala His Thr Leu Trp Ile Gly
260 265 270
Asn Ser Phe Met Leu Val Thr Ser Ala Gly Leu Met Ala His His Leu
275 280 285
Phe Gly Cys Trp His Gly Asp Arg Arg Leu Ser Ala Lys Tyr Gly Glu
290 295 300
Ala Phe Glu Ile Val Lys Ala Arg Thr Ser Thr Phe Pro Leu Gln Ala
305 310 315 320
Ile Trp Glu Gly Arg Gln Val Leu Pro Ala Asp Tyr Tyr Lys Glu Phe
325 330 335
Leu Arg Ala Pro Tyr Phe Ala Val Thr Ala Phe Thr Leu Gly Ala Tyr
340 345 350
Phe Ala His Pro Ile Met Gln Ser Ala Ser Phe Tyr Leu Arg Trp
355 360 365
<211> 1749
<212> DNA
<213> 人工序列(Artificial Sequence)
<220>
<223> 杜氏巴氏藻(Dunaliella bardawil):Zds
<400> 5
atgttggggc tgcagagcaa ggaatcgcaa ctgtgcacca caaatgtgcc agccaggcgt 60
ggctatgcgc agtgcaccag tacccgcacc cgcagacgga cgcgctgcac cacccaagcc 120
attgccaccc cccctgctcc tccaaagacc acacccaggg agtggaccac ccaggatgtg 180
agcaaggtgg ccttgaagga tgtgcctttg aagtccttgt acccggatga gcctgcccct 240
ccaaagccag gtgcccccaa gatgcgtgtg gccattgtgg gcagtggact ggctggcctg 300
tcgacagcag tggagctgct agaccaaggg catgaggtgg acatctatga ccagcgcagc 360
tgggttggag gcaaggtggc ctcatggcaa gacaaggatg gcaaccacat tgagatgggc 420
ttgcacgtct tcttcggctg ctaccacaac cttttccgtc tgatggccaa gtgtggagta 480
ctggagaact tgctggtgaa ggagcatact cacacctttt gcaacaatga tggggatgtc 540
agggagcttg acttccgttt tgaggtcgga ggacagaaaa tcggggctcc cttccatggc 600
ctgaaagcct tcttcacaac cccccagctt tctgtgggag acaaggcagc caatgcgctg 660
gcgctgggca ccagccctat tgtgcgctcc ttgatagacc cagagggggg catgagtgat 720
gtgcgaaacc tggacaacat cagcttctgg gactggttca agagccatgg cgggtctgag 780
caatccatga agcgcatgtg ggatcccatt gcgtatgctt tgggtttctt ggactgcaaa 840
gacatcagtg cgcgctgcat gctgaccatc ttccagttct ttgccaccaa gaccgacgcc 900
tccgtcctgc gcatgctgaa cggatctcca gctgagaggc tcctgaagcc catcaccgac 960
tacattgagg ccaagggagg ccgcatccac ttgcgtcagg gttgcaagga ggttctattt 1020
gaggatggcc ctgacggcaa gcctgtagtg accggcatgt gcatgggccg ggatggccaa 1080
gttgtgaagg ctgatgccta tgttgcagcc ctggacgtcc ctggggcgaa gcagctcttg 1140
ccgcaggcat ggagaaagta cccccagttt gacaaaatct acaagctcaa tggcgtgcct 1200
gtgatcactg tacagctgcg ctacaatggt tgggtgacgg agatgcagga cccagagaag 1260
gtcaagcagc tgacccagcc ccaaggcatc aacaacctgt tgtacagccc tgatgcgttc 1320
ttctcctgct tcgccgacct tgcccttgtg agccctgtgg agtacttcca tgagggcaag 1380
ggctccctca tgcaagtcgt gatcacgcct gctgccccgt acatgccatg gaccaacgag 1440
gccattgctg aggaggctga ccggcaggtc cgccagctct tccccagcgc ccgtaagctg 1500
gacatgatct ggcatagtgt tgtgaagatt ggccagtcct tgtaccagga ggccccaggc 1560
atggaccctt acaggcccga gcaggccacg cctgtgccca acttcttcct ggcaggcagc 1620
tacaccaagc aagactacat cgactcaatg gagggcgcca ccttgtcagg tcggcagtgt 1680
gctggtgaga ttatgaaggc tgtgcccttg atccaaagcc tgtcaaaggc gccacttccg 1740
agcatgtaa 1749
基于杜氏藻代谢途径的β-胡萝卜素高产工程菌及其构建方法与应用专利购买费用说明
![]()
Q:办理专利转让的流程及所需资料
A:专利权人变更需要办理著录项目变更手续,有代理机构的,变更手续应当由代理机构办理。
1:专利变更应当使用专利局统一制作的“著录项目变更申报书”提出。
2:按规定缴纳著录项目变更手续费。
3:同时提交相关证明文件原件。
4:专利权转移的,变更后的专利权人委托新专利代理机构的,应当提交变更后的全体专利申请人签字或者盖章的委托书。
Q:专利著录项目变更费用如何缴交
A:(1)直接到国家知识产权局受理大厅收费窗口缴纳,(2)通过代办处缴纳,(3)通过邮局或者银行汇款,更多缴纳方式
Q:专利转让变更,多久能出结果
A:著录项目变更请求书递交后,一般1-2个月左右就会收到通知,国家知识产权局会下达《转让手续合格通知书》。














动态评分
0.0