

IPC分类号 : C12N9/20I,C12N15/55I,C12N15/70I,C12N1/21I,C12P23/00I,C12R1/19N








专利摘要
本发明一种宏基因来源的脂肪酶、编码基因、载体、工程菌及在叶黄素制备中的应用,属于基因工程技术领域。宏基因来源的脂肪酶HsLIP1的氨基酸序列如SEQIDNO.1所示。编码脂肪酶HsLIP1的基因。基因的核苷酸序列如SEQIDNO.2所示。一种含有所述基因的重组载体。一种由所述重组载体转化得到的基因工程菌。所述基因在制备重组脂肪酶HsLIP1中的应用。所述的脂肪酶HsLIP1在叶黄素制备中的应用。本发明具有以下优点:脂肪酶HsLIP1易于原核表达,可以采用重组载体、菌株及制备方法高效生产本发明脂肪酶;而且,本发明酶的制备方法简便高效,易于大量表达,适于工业化生产。
权利要求
1.一种宏基因来源的脂肪酶HsLIP1,其特征在于:所述宏基因来源的脂肪酶HsLIP1 的氨基酸序列如SEQ ID NO.1所示。
2.一种编码权利要求1所述脂肪酶HsLIP1的基因。
3.如权利要求2所述的基因,其特征在于:所述基因的核苷酸序列如SEQ ID NO.2所示。
4.一种含有权利要求2或3所述基因的重组载体。
5.根据权利要求4所述的重组载体,其特征在于:所述重组载体是由编码HsLIP1的基因与pET-28a载体进行粘性互补连接得到。
6.一种由权利要求4或5所述重组载体转化得到的基因工程菌。
7.根据权利要求6所述的基因工程菌,其特征在于:所述基因工程菌是由重组载体转化至
8.权利要求2或3所述的基因在制备重组脂肪酶HsLIP1中的应用。
9.权利要求1所述的脂肪酶HsLIP1在叶黄素制备中的应用,其特征在于:脂肪酶HsLIP1在制备叶黄素中的应用是指脂肪酶HsLIP1在催化叶黄素浸膏底物的水解释放叶黄素的反应中的应用。
10.如权利要求9所述的脂肪酶HsLIP1在叶黄素制备中的应用,其特征在于:所述叶黄素浸膏底物为C12-C18长链脂肪酸酯化形成的单酯或双酯。
11.如权利要求10所述的脂肪酶HsLIP1在叶黄素制备中的应用,其特征在于:所述C12-C18长链脂肪酸为肉豆蔻酸、油酸、亚油酸或棕榈酸。
说明书
技术领域
本发明属于基因工程技术领域,具体涉及一种宏基因来源的脂肪酶、编码基因、载体、工程菌及在叶黄素制备中的应用。
背景技术
叶黄素属于类胡萝卜素,包含着多个共轭双键,这些共轭双键不仅赋予了颜色特征,也使其在疾病防治中具有较强的抗氧化功能,如阻止或预防像癌症、动脉硬化、白内障、色斑退化及其它类疾病。目前大规模生产而得到大量叶黄素晶体的方法是从天然资源中提取、分离和纯化得到。植物中的叶黄素类往往是与一些C12-C18长链脂肪酸如肉豆蔻酸、油酸、亚油酸和棕榈酸等酯化形成单酯或双酯的形式存在。目前运用最多的方法是利用强碱皂解叶黄素酯,再重结晶得到叶黄素。然而该方法有以下几个缺点:用到大量的强碱;过程中产生大量的废水;使用到一些有毒的有机溶剂;叶黄素的收率较低。
脂肪酶(triacylglycerol acylhydrolase,EC 3.1.1.3)是一类特殊的酯键水解酶,被定义为能够水解长链脂肪酸酯的酶,广泛存在于各种动物、植物和微生物(包括霉菌、酵母及细菌)中,微生物是目前工业脂肪酶的主要来源。利用脂肪酶在双相系统中(即油-水界面)能催化酯键的水解这一特性将叶黄素脂肪酸脂(叶黄素酯)水解释放叶黄素晶体。脂肪酶水解条件温和,效率高,最低限度的使用有机溶剂,精制步骤简洁,因此该酶在制备叶黄素晶体领域具备广阔的应用空间。
宏基因组学分析不需要对环境的所有微生物进行培养,它可以直接通过对环境细胞进行裂解,然后从基质和细胞碎片中分离回收DNA,以此分析微生物群落的组成和结构,使得原本超过99%的不可培养微生物基因资源的研究和开发成为了现实,极大限度地开发环境微生物资源的潜能。
发明内容
本发明的目的在于公开了一种宏基因来源的脂肪酶、编码基因、载体、工程菌及在叶黄素制备中的应用。
本发明的目的是通过以下技术方案实现的:
一种宏基因来源的脂肪酶HsLIP1,其中,所述宏基因来源的脂肪酶HsLIP1的氨基酸序列如SEQ ID NO.1所示。
一种编码上述技术方案所述脂肪酶HsLIP1的基因。
如上述技术方案所述的基因,其中,所述基因的核苷酸序列如SEQ ID NO.2所示。
一种含有上述技术方案所述基因的重组载体。
根据上述技术方案所述的重组载体,其中,所述重组载体是由编码HsLIP1的基因与pET-28a载体进行粘性互补连接得到。
一种由上述技术方案所述重组载体转化得到的基因工程菌。
根据上述技术方案所述的基因工程菌,其中,所述基因工程菌是由所述的重组质粒转化至E.coli BL21感受态细胞中得到。
上述技术方案所述的基因在制备重组脂肪酶HsLIP1中的应用。
上述技术方案所述的脂肪酶HsLIP1在叶黄素制备中的应用,其中,脂肪酶HsLIP1在制备叶黄素中的应用是指脂肪酶HsLIP1在催化叶黄素浸膏底物的水解释放叶黄素的反应中的应用。本技术方案中叶黄素浸膏底物含叶黄素脂肪酸酯。
如上述技术方案所述的脂肪酶HsLIP1在叶黄素制备中的应用,其中,所述叶黄素浸膏底物为C12-C18长链脂肪酸酯化形成的单酯或双酯。优选的所述C12-C18长链脂肪酸为肉豆蔻酸、油酸、亚油酸或棕榈酸。
本发明中宏基因来源的脂肪酶HsLIP1,其氨基酸序列如SEQ ID NO.1所示,即:HGFTGWGREEMFGFKYWGGVRGDIEQWLNDNGYRTYTLAVGPLSSNWDRACEAYAQLVGGTVDYGAAHAAKHGHARFGRTYPGLLPELKRGGRIHIIAHSQGGQTARMLVSLLENGSQEEREYAKAHNVSLSPLFEGGHHFVLSVTTIATPHDGTTLVNMVDFTDRFFDLQKAVLEAAAAASNVPYTSQVYDFKLDQWGLRRQPGESFDHYFERLKRSPVWTSTDTARYDLSVSGAEKLDQWVQASPNTYYLSFSTERTYRGALTGNHYPELGMNAFSAVVCAPFLGPYRNPTLGIDDRWLENDGIVNTVSMNGPKRGSSDRIVPYDGTL。
本发明中所述脂肪酶HsLIP1,具备pH耐受范围广且金属离子Ni2+、Na+和Mg2+以及有机溶剂二甲基亚砜(DMSO)和乙二醇对该脂肪酶的水解活性有一定促进作用等特征。
由于氨基酸序列的特殊性,任何含有SEQ ID NO.1所示氨基酸序列的肽蛋白的片段或其变体,如其保守性变体、生物活性片段或衍生物,只要该肽蛋白的片段或肽蛋白变体与前述氨基酸序列同源性在95%以上,均属于本发明保护范围之列。具体的所述改变可包括氨基酸序列中氨基酸的缺失、插入或替换;其中,对于变体的保守性改变,所替换的氨基酸具有与原氨基酸相似的结构或化学性质,如用亮氨酸替换异亮氨酸,变体也可具有非保守性改变,如用色氨酸替换甘氨酸。本发明所述肽蛋白的片段、衍生物或类似物是指基本上保持本发明所述的脂肪酶相同的生物学功能或活性的肽蛋白,可以是下列情形:(Ⅰ)一个或多个氨基酸残基被保守或非保守氨基酸残基(优选的是保守氨基酸残基)取代,并且取代的氨基酸可以是也可以不是由遗传密码子编码的;(Ⅱ)一个或多个氨基酸残基上的某个基团被其它基团取代;(Ⅲ)成熟肽蛋白与另一种化合物(比如延长肽蛋白半衰期的化合物,例如聚乙二醇)融合;(Ⅳ)附加的氨基酸序列融合进成熟的肽蛋白而形成的肽蛋白序列(如用来纯化此肽蛋白的序列或蛋白原序列)。
所述肽蛋白可以是重组蛋白、天然蛋白或合成蛋白,可以是纯天然纯化的产物,或是化学合成的产物,或使用重组技术从原核或真核宿主(例如:细菌、酵母、高等植物、昆虫和哺乳动物细胞)中产生。根据重组生产方案所用的宿主,本发明的肽蛋白可以是糖基化的。本发明的肽蛋白还可以包括或不包括起始的甲硫氨酸残基。
本发明还涉及编码所述脂肪酶HsLIP1的基因。
具体的,所述基因核苷酸序列可如SEQ ID NO.2所示,即:
CATGGGTTTA CCGGATGGGG ACGAGAGGAA ATGTTTGGAT TCAAGTATTG GGGCGGCGTGCGCGGCGATA TCGAACAATG GCTGAACGAC AACGGTTATC GAACGTATAC GCTGGCGGTC GGACCGCTCTCGAGCAACTG GGACCGGGCG TGTGAAGCGT ATGCTCAGCT TGTCGGCGGG ACGGTCGATT ATGGGGCAGCCCATGCGGCA AAGCACGGCC ATGCGCGGTT TGGCCGCACT TATCCCGGCC TGTTGCCGGA ATTGAAAAGGGGTGGCCGCA TCCATATCAT CGCCCACAGC CAAGGGGGGC AGACGGCCCG CATGCTTGTC TCGCTCCTAGAGAACGGAAG CCAAGAAGAG CGGGAGTACG CCAAGGCGCA TAACGTGTCG TTGTCACCGT TGTTTGAAGGTGGACATCAT TTTGTGTTGA GTGTGACGAC CATCGCCACT CCTCATGACG GGACGACGCT TGTCAACATGGTTGATTTCA CCGATCGCTT TTTTGACTTG CAAAAAGCGG TGTTGGAAGC GGCGGCTGCC GCCAGCAACGTGCCGTACAC GAGTCAAGTA TACGATTTTA AGCTCGACCA ATGGGGACTG CGCCGCCAGC CGGGTGAATCGTTCGACCAT TATTTTGAAC GGCTCAAGCG CTCCCCTGTT TGGACGTCCA CAGATACCGC CCGCTACGATTTATCCGTTT CCGGAGCTGA GAAGTTGGAT CAATGGGTGC AAGCAAGCCC GAATACGTAT TATTTGAGTTTCTCTACAGA ACGGACGTAT CGCGGAGCGC TCACAGGCAA CCATTATCCC GAACTCGGAA TGAATGCATTCAGCGCGGTC GTATGCGCTC CGTTTCTCGG TCCGTACCGC AATCCGACGC TCGGCATTGA CGACCGATGGTTGGAGAACG ATGGCATTGT CAATACGGTT TCCATGAACG GTCCAAAGCG TGGATCAAGC GATCGGATCGTGCCGTATGA CGGGACGTTG。
由于核苷酸序列的特殊性,任何SEQ ID NO.2所示多核苷酸的变体,只要其与该多核苷酸具有70%以上同源性,均属于本发明保护范围之列。所述多核苷酸的变体是指一种具有一个或多个核苷酸改变的多核苷酸序列。此多核苷酸的变体可以使生的变位变异体或非生的变异体,包括取代变异体、缺失变异体和插入变异体。如本领域所知的,等位变异体是一个多核苷酸的替换形式,它可能是一个多核苷酸的取代、缺失或插入,但不会从实质上改变其编码的肽蛋白的功能。
本发明的要点在于提供了SEQ ID NO.1所示的氨基酸序列和SEQ ID NO.2所示的核苷酸序列,在已知该氨基酸序列和核苷酸序列的情况下,该氨基酸序列和核苷酸序列的获得,以及相关载体、宿主细胞的获得,对于本领域技术人员来说均是显而易见的。
本发明还涉及含有所述基因的重组载体,以及由所述重组载体转化得到的基因工程菌。
本发明还涉及所述的基因在制备重组脂肪酶HsLIP1中的应用。具体的,所述应用为:构建含有所述编码基因的重组载体,将所述重组载体转入大肠杆菌中,获得的重组基因工程菌进行诱导培养,培养液分离得到含有重组脂肪酶的菌体细胞,菌体细胞经细胞碎、分离纯化获得所述重组脂肪酶HsLIP1。
经实验证明,本发明所述脂肪酶HsLIP1在宿主菌中表达产量高,例如在埃希氏大肠杆菌中过表达,HsLIP1的可溶性表达高。以对硝基苯酚脂为底物,在pH8.0,50℃条件下,HsLIP1的酶活力最高,最适条件下的酶活为9.37U/mL,在35℃的水浴中孵育480min之后保持87%的相对剩余活力。50ug的HsLIP1催化叶黄素浸膏(叶黄素脂肪酸酯)的水解反应30min释放0.537ug叶黄素,金属离子Ni2+、Na+及Mg2+以及有机溶剂二甲基亚砜(DMSO)和乙二醇对HsLIP1的水解活性有一定促进作用。
本发明的脂肪酶HsLIP1具有高表达量,高活性,底物特异性,同时本发明脂肪酶HsLIP1可以催化叶黄素浸膏中酯键水解释放出叶黄素和相应的C12-C18长链脂肪酸如如肉豆蔻酸、油酸、亚油酸和棕榈酸等。
本发明的高表达、高活性的脂肪酶HsLIP1的制备方法及酶学表征:
(1)、本发明所述脂肪酶HsLIP1基因全长的获取。设计脂肪酶简并引物,以本实验室采集的土壤(水)样品包括宁波、洞头、象山、天台、太湖水、浙江新昌、西安附近黄土高原、深圳浅水湾红土等八个不同地区的土壤(水)样品提取的宏基因组DNA为模板,克隆获得宏基因组新酶1条,得到全长脂肪酶基因,即脂肪酶HsLIP1,SEQIDNO.2所示的脂肪酶核苷酸全长序列。
(2)、含目的基因的表达载体系统的构建。将步骤(1)中所述的脂肪酶基因克隆至表达载体,如pET-28a。
(3)、将步骤(2)中含脂肪酶HsLIP1基因的重组载体转入异源表达宿主细胞中,如(Escherichiacoli)BL21,在适合表达的条件下,培养重组宿主细胞。
(4)、从步骤(3)的培养物中分离纯化出本发明中所述的高表达、高活性的脂肪酶HsLIP1。
(5)、通过如上制备方法,对获得的脂肪酶进行进一步酶学特征表征,包括最适温度、热稳定性、最适pH值、重金属离子溶剂以及有机溶剂的耐受性等。
本发明还涉及所述的脂肪酶HsLIP1在催化叶黄素浸膏的水解反应释放叶黄素的应用。
所述叶黄素浸膏底物包含一些C12-C18长链脂肪酸如肉豆蔻酸、油酸、亚油酸和棕榈酸等酯化形成的单酯或双酯。
本发明依据数据库中的脂肪酶序列,设计了保守位点引物,进而在宏基因组中扩增出了同源片段,构建了包含宏基因组同源片段的新酶(HsLIP1),对其进行了功能表征,并将其应用到脂水解反应中。以对硝基苯酚脂作为底物,该酶在pH8.0,50℃条件下的酶活力最高,最适条件下的酶活为9.37U/mL,在35℃的水浴中孵育480min之后保持87%的相对剩余活力。50ug的HsLIP1催化叶黄素浸膏的水解反应30min释放0.537ug叶黄素,金属离子Ni2+、Na+和Mg2+以及有机溶剂二甲基亚砜(DMSO)和乙二醇对该脂肪酶HsLIP1的水解活性有一定促进作用。
本发明具有以下有益效果:
本发明的脂肪酶HsLIP1易于原核表达,可以采用本专利中所述的重组载体、菌株及制备方法高效生产本发明脂肪酶;而且,本发明酶的制备方法简便高效,易于大量表达,适于工业化生产。
附图说明:
1、图1为宏基因组DNA的琼脂糖凝胶电泳图。
2、图2为宏基因组PCR产物的琼脂糖凝胶电泳图。
3、图3为菌落鉴定PCR产物的琼脂糖凝胶电泳图。
4、图4为脂肪酶诱导表达结果SDS-PAGE电泳图;
5、图5为脂肪酶纯化结果SDS-PAGE电泳图;
6、图6为对硝基苯酚标准曲线;
7、图7为脂肪酶不同温度反应;
8、图8为脂肪酶热稳定性反应;
9、图9为脂肪酶不同pH值反应;
10、图10为叶黄素标准曲线;
11、图11为脂肪酶对叶黄素浸膏的不同温度反应;
12、图12为脂肪酶对叶黄素浸膏的不同pH值反应;
13、图13为脂肪酶对叶黄素浸膏的不同金属离子反应;
14、图14为脂肪酶对叶黄素浸膏的不同有机溶剂反应;
具体实施方式:
为使本发明的技术方案便于理解,下面结合具体实施例对本发明进行进一步描述,但本发明的保护范围并不仅限于此:
本发明中的实验方法如无特别说明均为常规方法,具体可参见“MolecularCloning:ALaboratoryManual”(SambrookandRussell,ed.2001)。
本发明的实施例中使用的大肠杆菌DH5α、BL21(DE3)购自TransGen Biotech公司;引物合成由上海捷瑞生物工程技术有限公司完成;序列测序工作由生工生物工程(上海)股份有限公司完成。
实施例1:脂肪酶HsLIP1的获得以及含脂肪酶HsLIP1的重组菌的构建:
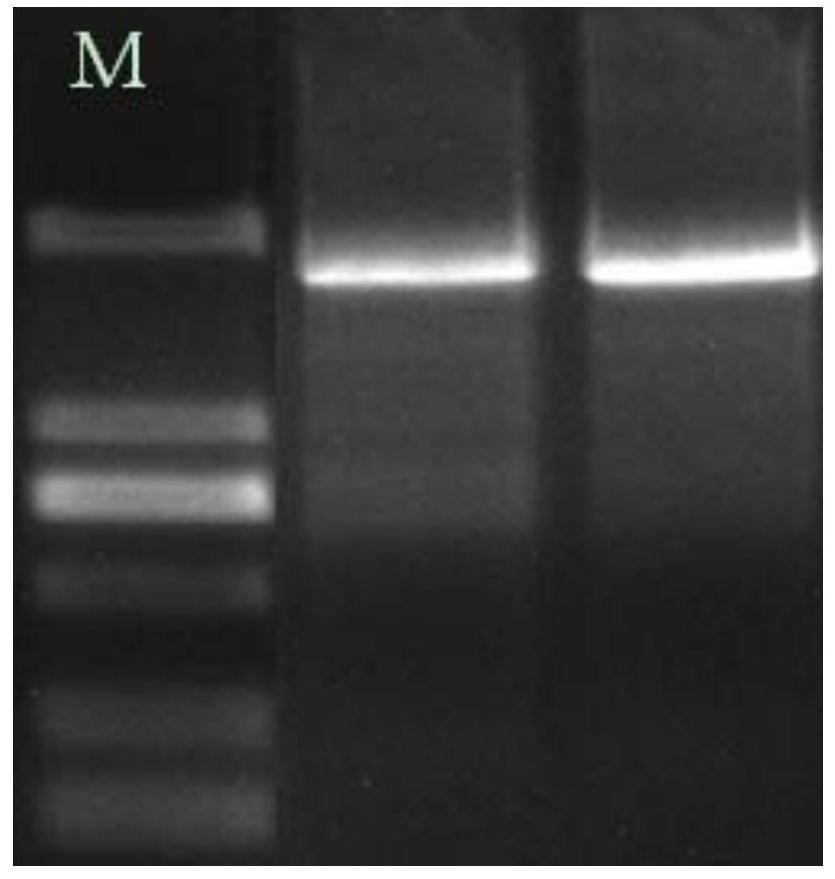
1、从中国各地不同地域的野外环境包括宁波、洞头、象山、天台、太湖水、浙江新昌、西安附近黄土高原、深圳浅水湾红土等八个不同地区,共采集10个土壤(水)样品,其中3号土壤样本来源于象山土壤。土壤组分复杂,提取的宏基因组DNA含有棕黑色或黑色的腐殖酸等杂质,它们对PCR等后续反应有极大的抑制作用,因此需要有效地对土壤中腐殖酸进行去除。采用Humic acid-Be-Gone A试剂盒有效去除土壤样品腐殖酸,回收沉淀的土壤可直接用于DNA提取。采用Soil Genomic DNAKit土壤基因组DNA提取试剂盒提取土壤样品基因组DNA,收集的DNA溶液,琼脂糖凝胶电泳检测,电泳图如图1所示,列M:15,000DNA marker;列M’:10,000DNA marker;列1-10:Metagenomic DNA(宏基因组DNA);并使用琼脂糖凝胶DNA回收试剂盒(Generay,上海)回收DNA,于-20℃保存。
2、依据来源于G.thermocatenulatus的活性较高的脂肪酶基因BTL2(GenBank编号为ACCESSIONX95309)和来源于Geobacillus stearothermophilus的脂肪酶基因菌T6(EMBL登录号AF429311.1),进行序列比对分析查找保守区序列,设计引物。
表1本发明PCR扩增引物序列
以宏基因组DNA为模板,用高保真酶进行PCR扩增,配制反应体系如下:
上述体系混匀后,设置程序如下:
PCR扩增结束后,将PCR产物进行琼脂糖凝胶电泳检测,检测时稳压稳流电泳仪设置的电压120V,电流为120mA,时间为30min;琼脂糖凝胶电泳图如图2所示,其中M为marker、列1为PCR产物,目的产物大小为1000bp左右,将正确位置的含有目的产物胶切下并回收,得到SEQ ID NO.2的核苷酸序列,具体操作步骤详见GENEray薄型琼脂糖凝胶DNA回收试剂盒(Generay,上海)说明书。
3、将上一步纯化回收的目的产物DNA连接至pUCm-T载体,反应体系如下:
按上述体系混匀后置于16℃恒温水浴连接过夜。
根据CaCl2法制备E.coli DH5α感受态细胞,将连接过夜的重组质粒转化至DH5α感受态细胞中,具体操作步骤如下:
(1)向-80℃冰箱取出的100μLDH5α感受态细胞(冰上解冻)加入10μL连接产物,冰浴30min;
(2)42℃热激90s;
(3)迅速放在冰上,继续冰浴2min;
(4)加入500μL无抗性的LB液体培养基,37℃,200rpm,摇菌1h;
(5)取100μL菌液均匀涂布在含0.1mM IPTG和20μg/mL X-gal的蓝白斑筛选的Amp抗性的LB固体平板上,先将平板正向放置,37℃培养1h,后倒置过夜培养。
培养16h后,LB固体平板上长出单个菌落,挑取LB固体平板上的白色单克隆菌落,加到10μL无菌水中,取2μL进行PCR阳性鉴定,菌落PCR反应体系如下:
上述体系混匀后,设置PCR反应程序如下:
取3μLPCR扩增产物进行琼脂糖凝胶电泳,验证目的DNA片段是否已经插入,检测插入片段的大小,并且挑选合适大小的片段克隆送到基因测序公司(北京擎科新业生物技术有限公司杭州测序部)进行测序,菌落鉴定PCR产物的琼脂糖凝胶电泳图如图3所示,其中M为marker、列1~列3为PCR产物;测序引物为M13:
M13F:CGCCAGGGTTTTCCCAGTCACGAC
M13R:CACACAGGAAACAGCTATGAC。
通过前面的操作,我们已经得到了野生型脂肪酶基因(MetlipXS)全长,命名为MetlipXS,鉴定测序,即得到序列为SEQ ID NO.1所示脂肪酶重组菌的全长序列。
剩余的8μL菌液加到1mL含Amp的LB培养基中培养,37℃,200rpm,过夜培养,用北京全式金质粒提取试剂盒EasyPure PlasmidMiniPrep Kit分别提取培养好的重组质粒即含有HsLIP1目标DNA的pUCm-T质粒。
4、使用EcoR I和Hind III酶将pET-28a质粒和含有HsLIP1目标DNA的pUCm-T质粒进行双酶切以产生对应互补的黏性末端。酶切体系如下所示:
按上述体系混匀后置于37℃恒温酶切反应15min,将酶切产物进行琼脂糖凝胶电泳检测。在正确位置的胶切下并回收,具体操作步骤详见薄型琼脂糖凝胶DNA回收试剂盒(Generay,上海)说明书。
将经同样酶切回收的HsLIP1基因与pET-28a载体进行粘性互补连接,连接体系如下:
按上述体系混匀后置于25℃恒温水浴连接反应20min,得到HsLIP1基因与pET-28a载体的重组质粒。
5、将重组质粒转化至E.coli BL21感受态细胞中,将菌液均匀涂布在含0.1mMIPTG含有乳化不透明的甘油酯底物筛选的卡那霉素Kan抗性的LB固体平板上,先将平板正向放置,37℃培养1h,以吸收过多的液体,倒置过夜培养,得到具有透明圈的菌落即为重组菌E.coli BL21/pET28a-HsLIP1。
挑取具有透明圈的菌落进行菌落PCR鉴定,引物为T7。菌落PCR的方法同含有HsLIP1目标DNA的pUCm-T质粒的阳性鉴定。选取合适大小克隆片段送上海生工测序,测序引物为T7。所述引物购自北京擎科新业生物技术有限公司杭州测序部,T7序列为:
T7:TAATACGACTCACTATAGGG
T7t:GCTAGTTATTGCTCAGCGG
至此,脂肪酶HsLIP1的大肠杆菌异源表达体系即E.coliBL21/pET28a/HsLIP1脂肪酶表达系统构建完成。
实施例2:重组菌的诱导表达与脂肪酶HsLIP1的纯化:
按1:100的比例接种重组菌E.coli BL21/pET28a-HsLIP1至100mL含卡那霉素(50mg/mL)的LB液体培养基,37℃,200rpm培养,摇床振荡培养至OD600=0.6,加入IPTG至终浓度为0.4mM进行诱导,另外一组不加IPTG作为未诱导对照,25℃,180rpm诱导表达14h。将过夜诱导的菌液在4℃条件下,6000rpm离心5min,弃上清。用10mL PBS(pH 7.4)缓冲液将菌重悬体,同等条件下继续离心5min,弃上清收集菌体。加10mL含有2mMβ-巯基乙醇的PBS(pH7.4)重悬菌体。将重悬混合液置于冰水浴中,利用超声破碎仪进行细胞破碎25min。超声破碎仪参数设置如下:超声功率300W,超声时间3s,间隔时间3s。破碎完成后在4℃、12000rpm条件下离心15min,收集的上清即为粗酶液。
将收集的粗酶液过0.45μm的水系滤膜过滤后,用Ni-NTA纯化树脂进行纯化回收,获得本发明中所述的脂肪酶HsLIP1纯酶;镍柱亲和层析纯化目的蛋白的整个过程在4℃层析柜中进行,具体步骤如下:
(1)使用10倍体积的20mM Tris-HCl缓冲液(pH 7.4)平衡镍柱1h后放掉;
(2)将过滤好的粗酶液上样于柱中,结合1h后放掉结合柱里面的结合后的酶液;
(3)先用上样缓冲液洗柱,再用20倍体积的20mM咪唑缓冲液洗去杂蛋白,最后用10倍的250mM咪唑缓冲液将目的蛋白洗脱,收集洗脱液。
在4℃冰箱中进行透析,将最后收集的洗脱液放在透析袋中,置于20mM Tris-HCl缓冲液(pH 7.4)中,磁力搅拌6h后重新换新的缓冲液,重复3次;收集透析袋中酶液,即为纯化的脂肪酶酶液。对蛋白纯化的样品进行SDS-PAGE蛋白电泳检测,分析脂肪酶蛋白的纯化情况。
纯化过程用到的缓冲液:
1)上样缓冲液:Tris 20Mm(Ph7.4),NaCl 250mM;
2)20mM咪唑缓冲液:在上样缓冲液中加入咪唑至20mM;
3)250mM咪唑缓冲:在上样缓冲液中加入咪唑至250mM;
重组菌经IPTG诱导表达,破碎离心之后,经SDS-PAGE蛋白电泳分析,在40kD左右有一条明显的蛋白条带,与预期的目的条带吻合,脂肪酶诱导表达结果SDS-PAGE电泳图如图4所示,其中M:蛋白质低分子量标记;1:未诱导菌液;2:诱导菌液;3:诱导菌液上清。该脂肪酶蛋白表达量比较高,HsLIP1的可溶性表达高。蛋白纯化结果如图5所示,其中M:蛋白质低分子量标记;1:样品流出液;2:上样流出液;3:20mM咪唑洗脱液;4:250mM咪唑洗脱液。
实施例3:采用比色测定法中的对硝基苯酚法对脂肪酶的活力进行测定:
以p-NPP(对硝基苯酚棕榈酸酯)作为底物,脂肪酶能够水解对硝基苯酚棕榈酸酯,生成带有颜色的对硝基苯酚,对硝基苯酚在410nm处有特征吸收峰。吸光度与对硝基苯酚量呈良好的线性关系,因此可以根据反应液在410nm处的吸光度计算脂肪酶活力。
反应机理如图所示:
式I为脂肪酶水解对硝基苯酚棕榈酸酯的反应方程式
以p-NPP为底物测定脂肪酶活力的具体操作方法如下:
对硝基苯酚溶液(6mM):称取0.0835g对硝基苯酚(p-NP),先用1mL的95%乙醇溶解,再用蒸馏水定容至100mL。
硝基苯酚棕榈酸酯溶液:称取30mg对硝基苯酚棕榈酸酯(p-NPP),用异丙醇定容至10mL。
(1)标准曲线的绘制:向离心管中分别加入p-NP溶液10μL、15μL、20μL、30μL、40μL、50μL,分别加入50mM Tris-HCl(pH 8.0)缓冲液至2mL。然后每管加入0.5mL 10%三氯乙酸溶液和0.5mL 10%碳酸钠溶液,摇晃使其混合均匀,使用分光光度计于410nm处测定吸光值,绘制酶活性标准曲线.
(2)测定脂肪酶酶活:向15mL的试管中加入0.1mLp-NPP底物溶液和1.8mL 50mMTris-HCl(pH 8.0)缓冲液,50℃恒温水浴5min。实验组加入0.1mL酶液,立即摇晃使其混合均匀,在50℃恒温水浴10min后立即加入5mL 0.5M三氯乙酸溶液终止反应,再加入5.25mL0.5M氢氧化钠溶液显色,在410nm波长下测定酶催化产生的对硝基苯酚的吸收值(空白组加入0.1mL去离子水代替0.1mL酶液)。
脂肪酶对p-NPP的酶活定义:在50℃,pH 8.0条件下,每1min释放1μmol对硝基苯酚所需的酶量。
酶活计算公式:
U为样品酶活(U/mL),C为p-NP浓度(μmol/mL),n为稀释倍数,V为反应液的体积(mL),V'为酶液的体积(mL),T为反应时间(min)。
结果表明,对硝基苯酚浓度与吸光值呈良好的线性关系,如图6,对硝基苯酚标准曲线为:y=0.1553x+0.0209,R2=0.9985
在410nm处测得的吸光值A410为1.218,根据标准曲线公式计算得脂肪酶的比活为9.37U/mL。
实施例4:脂肪酶HsLIP1的最适反应温度和热稳定性分析
以pH 8.0的PBS为缓冲液,分别在30-80℃的范围内以10℃的间隔在水浴中反应10min,紫外分光光度计410nm波长下测定动力学曲线。将纯化脂肪酶样品分别40℃、50℃和60℃条件下孵育30、60、90、120、150、180和240min后,在最适条件下以p-NPP为底物,测定脂肪酶残留活性,分析温度对脂肪酶稳定性的影响。
脂肪酶不同温度值反应如图7所示。脂肪酶在pH 7.4的缓冲液中的最适反应温度是50℃,在40-60℃的温度范围内,脂肪酶的活力保持在80%以上,在80℃的条件下,依然保持46%的酶活力。为测定脂肪酶的热稳定性,将纯酶液孵育在pH 8.0的缓冲液中,不同时间测定它们的相对剩余活性,结果如图8所示,在50℃处理180min后,脂肪酶的活力剩余80%以上,在65℃的条件下,脂肪酶活力下降的比较快,处理240min后,脂肪酶的活力只有20%的残余活力,而在35℃条件下,脂肪酶的活性几乎没有影响。
实施例5:脂肪酶HsLIP1的最适反应pH值分析
为测定脂肪酶最适pH,配制pH 5.0-10.0的50mM缓冲液如下:Glycine-HCl缓冲液(pH 5.0-6.0)、Tris-HCl/NaOH缓冲液(pH 7.0-8.0)、Tris-HCl缓冲液(pH 9.0)、Glycine-NaOH缓冲液(pH 10.0)。分别在不同pH缓冲液中,按照实施例4的体系,在37℃水浴中反应10min,紫外分光光度计410nm波长下测定脂肪酶对底物p-NPP的酶促反应动力学曲线。
不同pH值对酶促反应的影响如图9所示,脂肪酶在pH值为8.0缓冲液中的酶活最高,在pH值为10.0的缓冲液中,仍然保持55%的活力,当缓冲液的pH值在7.0及以下的时候,脂肪酶的活力开始迅速的下降,在pH值为5.0的条件下,相对活力只有23%。
实施例6:叶黄素标准曲线绘制
精确称取10mg叶黄素标准品溶于10ml的三氯甲烷,-80℃保存(两周内使用)。取1ml叶黄素三氯甲烷溶液溶于10ml正丙醇,形成0.1mg/ml的叶黄素正丙醇溶液。分别取10、20、30、40、50、100、200、300、400ul叶黄素正丙醇溶液定容于1000ul正己烷。
HPLC的仪器分析条件如下:
色谱柱:Agilent HC-C18柱(4.6mm×250mm,5μm);
流动相:乙腈,0.45μm滤膜过滤,超声脱气15min;
检测波长:446nm;
流速:0.6ml/min;
柱温:28℃;
进样量:20μl。
按制备的标准溶液系列上机测定,以峰面积为纵坐标(Y),浓度为横坐标(X)绘制标准曲线,见图10。叶黄素浓度与峰面积呈良好的线性关系,回归方程为:Y=77680X–35777,R2=0.9978。
实施例7:脂肪酶HsLIP1催化叶黄素浸膏的最适反应温度分析
叶黄素浸膏溶液:称取10mg叶黄素浸膏(总叶黄素含量15.8%),与8mL正丙醇混合,升温至65℃,搅拌直至形成一种均一流动的浸膏溶液。
为测定脂肪酶对叶黄素浸膏的最适反应温度,在Tris缓冲液(pH 7.4)中分别在30-80℃的范围内以10℃的间隔下测定脂肪酶对底物叶黄素浸膏的酶促反应动力学曲线。分别向EP管中加入350μl Tris缓冲液(pH 7.4),再加50μl浸膏溶液混匀,最后加100μl酶,混匀,分别在30、40、50、60、70、80℃下恒温水浴30min后,立即加入1ml正己烷,漩涡混合10s,离心1min,回收上层有机相1ml左右。用0.45μm的针筒式有机滤膜过滤器过滤后上机测定,HPLC的方法同实施例6。
脂肪酶不同温度值反应如图11所示。HsLIP1的最适温度是50℃。在pH值在40-50℃的条件下,HsLIP1表现出较好的活性。
实施例8:脂肪酶HsLIP1催化叶黄素浸膏的最适反应pH值分析
分别在pH 5.0-11.0缓冲液中,按照方法实施例7,在温度为50℃条件下,测定不同pH下脂肪酶对底物叶黄素浸膏的酶促反应动力学曲线。用0.45μm的针筒式有机滤膜过滤器过滤后上机测定。
脂肪酶不同温度反应如图12所示。HsLIP1的最适pH是8.0,pH值在7.0-9.0的范围内,HsLIP1表现出较好的活性。我们规定以温度为50℃,pH值为8.0条件下测得的脂肪酶活性为100。
实施例9:不同金属离子对脂肪酶HsLIP1催化叶黄素浸膏活性的影响
分别含有10mM不同金属离子LiCl、KCl、NaCl、CuCl2、CaCl2、BaCl2、FeSO4、MgSO4、NiSO4、AlCl3缓冲液中,在Tris缓冲液(pH 8.0)中50℃下按照实施例7的体系,测定在不同金属离子下脂肪酶对底物叶黄素浸膏的酶促反应动力学曲线。用0.45μm的针筒式有机滤膜过滤器过滤后上机测定。
不同金属离子对脂肪酶水解活性影响如图13所示。在10mM金属离子存在的条件下,其中Na+、Ni+、Ba2+、Mg2+及Ca2+对脂肪酶HsLIP1催化叶黄素浸膏的水解活性有促进作用,其中Ni2+对肪酶HsLIP1对叶黄素酯的水解活性的促进作用最明显,叶黄素的产量提高了151%。我们规定在没有金属离子存在,温度为50℃,pH值为8.0的条件下测得的脂肪酶活性为100。
实施例10:不同有机溶剂对脂肪酶HsLIP1活性的影响
分别含有25%不同有机溶剂缓冲液中,在Tris缓冲液(pH 8.0)中50℃下按照实施例7的体系,测定在不同有机溶剂下脂肪酶对底物叶黄素浸膏的酶促反应动力学曲线。用0.45μm的针筒式有机滤膜过滤器过滤后上机测定。
不同有机溶剂对脂肪酶水解活性影响如图14所示。在25%有机溶剂存在的条件下,脂肪酶对不同有机溶剂的耐受力都挺高,而DMSO和乙二醇对脂肪酶HsLIP1催化叶黄素酯的水解活性有一定促进作用。我们规定在没有有机溶剂存在,温度为50℃、pH值为8.0的条件下测得的脂肪酶活性为100。
以上所述,仅为本发明的较佳实施例,并非对本发明作任何形式上和实质上的限制,凡熟悉本专业的技术人员,在不脱离本发明技术方案范围内,当可利用以上所揭示的技术内容,而作出的些许更动、修饰与演变的等同变化,均为本发明的等效实施例;同时,凡依据本发明的实质技术对以上实施例所作的任何等同变化的更动、修饰与演变,均仍属于本发明的技术方案的范围内。
序列表
<110> 杭州师范大学
<120> 一种宏基因来源的脂肪酶、编码基因、载体、工程菌及在叶黄素制备中的应用
<160> 8
<170> SIPOSequenceListing 1.0
<210> 1
<211> 330
<212> PRT
<213> triacylglycerol acylhydrolase
<400> 1
His Gly Phe Thr Gly Trp Gly Arg Glu Glu Met Phe Gly Phe Lys Tyr
1 5 10 15
Trp Gly Gly Val Arg Gly Asp Ile Glu Gln Trp Leu Asn Asp Asn Gly
20 25 30
Tyr Arg Thr Tyr Thr Leu Ala Val Gly Pro Leu Ser Ser Asn Trp Asp
35 40 45
Arg Ala Cys Glu Ala Tyr Ala Gln Leu Val Gly Gly Thr Val Asp Tyr
50 55 60
Gly Ala Ala His Ala Ala Lys His Gly His Ala Arg Phe Gly Arg Thr
65 70 75 80
Tyr Pro Gly Leu Leu Pro Glu Leu Lys Arg Gly Gly Arg Ile His Ile
85 90 95
Ile Ala His Ser Gln Gly Gly Gln Thr Ala Arg Met Leu Val Ser Leu
100 105 110
Leu Glu Asn Gly Ser Gln Glu Glu Arg Glu Tyr Ala Lys Ala His Asn
115 120 125
Val Ser Leu Ser Pro Leu Phe Glu Gly Gly His His Phe Val Leu Ser
130 135 140
Val Thr Thr Ile Ala Thr Pro His Asp Gly Thr Thr Leu Val Asn Met
145 150 155 160
Val Asp Phe Thr Asp Arg Phe Phe Asp Leu Gln Lys Ala Val Leu Glu
165 170 175
Ala Ala Ala Ala Ala Ser Asn Val Pro Tyr Thr Ser Gln Val Tyr Asp
180 185 190
Phe Lys Leu Asp Gln Trp Gly Leu Arg Arg Gln Pro Gly Glu Ser Phe
195 200 205
Asp His Tyr Phe Glu Arg Leu Lys Arg Ser Pro Val Trp Thr Ser Thr
210 215 220
Asp Thr Ala Arg Tyr Asp Leu Ser Val Ser Gly Ala Glu Lys Leu Asp
225 230 235 240
Gln Trp Val Gln Ala Ser Pro Asn Thr Tyr Tyr Leu Ser Phe Ser Thr
245 250 255
Glu Arg Thr Tyr Arg Gly Ala Leu Thr Gly Asn His Tyr Pro Glu Leu
260 265 270
Gly Met Asn Ala Phe Ser Ala Val Val Cys Ala Pro Phe Leu Gly Pro
275 280 285
Tyr Arg Asn Pro Thr Leu Gly Ile Asp Asp Arg Trp Leu Glu Asn Asp
290 295 300
Gly Ile Val Asn Thr Val Ser Met Asn Gly Pro Lys Arg Gly Ser Ser
305 310 315 320
Asp Arg Ile Val Pro Tyr Asp Gly Thr Leu
325 330
<210> 2
<211> 990
<212> DNA
<213> triacylglycerol acylhydrolase
<220>
<221> misc_feature
<222> (1)..(990)
<400> 2
catgggttta ccggatgggg acgagaggaa atgtttggat tcaagtattg gggcggcgtg 60
cgcggcgata tcgaacaatg gctgaacgac aacggttatc gaacgtatac gctggcggtc 120
ggaccgctct cgagcaactg ggaccgggcg tgtgaagcgt atgctcagct tgtcggcggg 180
acggtcgatt atggggcagc ccatgcggca aagcacggcc atgcgcggtt tggccgcact 240
tatcccggcc tgttgccgga attgaaaagg ggtggccgca tccatatcat cgcccacagc 300
caaggggggc agacggcccg catgcttgtc tcgctcctag agaacggaag ccaagaagag 360
cgggagtacg ccaaggcgca taacgtgtcg ttgtcaccgt tgtttgaagg tggacatcat 420
tttgtgttga gtgtgacgac catcgccact cctcatgacg ggacgacgct tgtcaacatg 480
gttgatttca ccgatcgctt ttttgacttg caaaaagcgg tgttggaagc ggcggctgcc 540
gccagcaacg tgccgtacac gagtcaagta tacgatttta agctcgacca atggggactg 600
cgccgccagc cgggtgaatc gttcgaccat tattttgaac ggctcaagcg ctcccctgtt 660
tggacgtcca cagataccgc ccgctacgat ttatccgttt ccggagctga gaagttggat 720
caatgggtgc aagcaagccc gaatacgtat tatttgagtt tctctacaga acggacgtat 780
cgcggagcgc tcacaggcaa ccattatccc gaactcggaa tgaatgcatt cagcgcggtc 840
gtatgcgctc cgtttctcgg tccgtaccgc aatccgacgc tcggcattga cgaccgatgg 900
ttggagaacg atggcattgt caatacggtt tccatgaacg gtccaaagcg tggatcaagc 960
gatcggatcg tgccgtatga cgggacgttg 990
<210> 3
<211> 24
<212> DNA
<213> triacylglycerol acylhydrolase
<220>
<221> misc_feature
<222> (1)..(24)
<400> 3
catgggttta ccggatgggg acga 24
<210> 4
<211> 24
<212> DNA
<213> triacylglycerol acylhydrolase
<220>
<221> misc_feature
<222> (1)..(24)
<400> 4
caacgtcccg tcatacggca cgat 24
<210> 5
<211> 24
<212> DNA
<213> triacylglycerol acylhydrolase
<220>
<221> misc_feature
<222> (1)..(24)
<400> 5
cgccagggtt ttcccagtca cgac 24
<210> 6
<211> 21
<212> DNA
<213> triacylglycerol acylhydrolase
<220>
<221> misc_feature
<222> (1)..(21)
<400> 6
cacacaggaa acagctatga c 21
<210> 7
<211> 20
<212> DNA
<213> triacylglycerol acylhydrolase
<220>
<221> misc_feature
<222> (1)..(20)
<400> 7
taatacgact cactataggg 20
<210> 8
<211> 19
<212> DNA
<213> triacylglycerol acylhydrolase
<220>
<221> misc_feature
<222> (1)..(19)
<400> 8
gctagttatt gctcagcgg 19
一种宏基因来源的脂肪酶、编码基因、载体、工程菌及在叶黄素制备中的应用专利购买费用说明
![]()
Q:办理专利转让的流程及所需资料
A:专利权人变更需要办理著录项目变更手续,有代理机构的,变更手续应当由代理机构办理。
1:专利变更应当使用专利局统一制作的“著录项目变更申报书”提出。
2:按规定缴纳著录项目变更手续费。
3:同时提交相关证明文件原件。
4:专利权转移的,变更后的专利权人委托新专利代理机构的,应当提交变更后的全体专利申请人签字或者盖章的委托书。
Q:专利著录项目变更费用如何缴交
A:(1)直接到国家知识产权局受理大厅收费窗口缴纳,(2)通过代办处缴纳,(3)通过邮局或者银行汇款,更多缴纳方式
Q:专利转让变更,多久能出结果
A:著录项目变更请求书递交后,一般1-2个月左右就会收到通知,国家知识产权局会下达《转让手续合格通知书》。














动态评分
0.0