

IPC分类号 : C12N9/20I,C12N15/55I,C12N1/21I,C12P41/00I,C12P7/62I,C12R1/19N




专利摘要
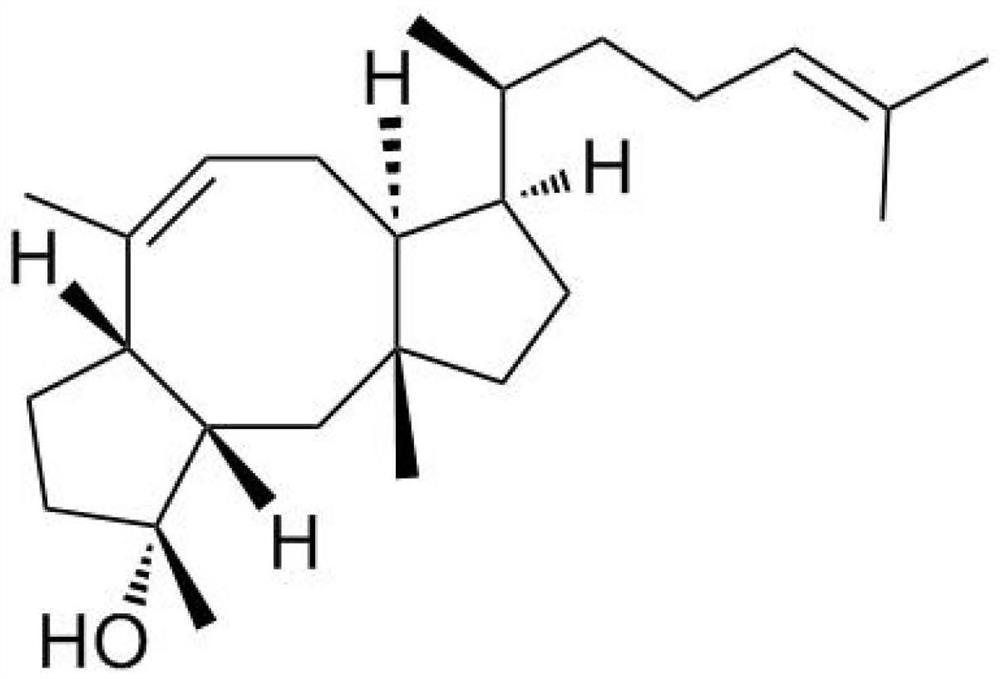
本发明提供了一种米曲霉脂肪酶及在拆分制备布瓦西坦中间体的应用,所述米曲霉脂肪酶的氨基酸序列如SEQIDNO.1所示,以发酵培养获得的湿菌体经冷冻干燥后得到的干菌体为催化剂,外消旋2‑(2‑叔丁氧基‑2‑氧代乙基)戊酸甲酯为底物,以pH7.0磷酸缓冲溶液为反应介质,在20‑60℃、1000rpm条件下进行拆分制备布瓦西坦中间体R‑2‑(2‑叔丁氧基‑2‑氧代乙基)戊酸,质量收率达到49.5%,产物R‑2‑(2‑叔丁氧基‑2‑氧代乙基)戊酸经甲酯化后对映体过量值>99%。
权利要求
1.一种米曲霉脂肪酶在拆分外消旋2-(2-叔丁氧基-2-氧代乙基)戊酸甲酯制备布瓦西坦手性中间体中的应用,其特征在于所述脂肪酶的氨基酸序列如SEQ ID NO.1所示。
2.如权利要求1所述的应用,其特征在于所述应用的方法为:以含米曲霉脂肪酶编码基因的工程菌经发酵培养获得的湿菌体冻干后的脂肪酶粗酶粉为生物催化剂,以外消旋2-(2-叔丁氧基-2-氧代乙基)戊酸甲酯为底物,以pH7.0缓冲液为反应介质,在20-60℃、1000rpm条件下进行拆分反应,反应完全后,将反应液分离纯化,获得布瓦西坦中间体R-2-(2-叔丁氧基-2-氧代乙基)戊酸。
3.如权利要求2所述的应用,其特征在于所述编码基因的核苷酸序列如SEQ ID NO.2所示。
4.如权利要求2所述的应用,其特征在于所述催化剂用量以缓冲液体积计为20g/L-100g/L,所述底物体积终浓度为1%-3%。
5.如权利要求2所述的应用,其特征在于反应时间为5min-180min。
6.如权利要求2所述的应用,其特征在于所述缓冲液为pH 7.0,0.2mM的Na
7.如权利要求2所述的应用,其特征在于所述催化剂按如下方法制备:将含米曲霉脂肪酶编码基因的工程菌接种在LB培养基中,37℃培养OD
8.如权利要求2所述的应用,其特征在于所述反应液分离纯化制备产物的方法为:反应结束后,反应液先用的2M NaOH水溶液调pH至9.0,加入等体积的环己烷萃取分离未反应的S-2-(2-叔丁氧基-2-氧代乙基)戊酸甲酯;然后用4M HCl水溶液酸化至pH 2.0,加入等体积乙酸乙酯萃取,分液漏斗分离出有机相,再经纯水洗涤两次,饱和NaCl洗涤两次,干燥,得到R-2-(2-叔丁氧基-2-氧代乙基)戊酸。
说明书
(一)技术领域
本发明属于生物酶催化领域,涉及一种脂肪酶立体选择性催化水解外消旋2-(2-叔丁氧基-2-氧代乙基)戊酸甲酯对映体制备布瓦西坦中间体R-2-(2-叔丁氧基-2-氧代乙基)戊酸的应用。
(二)背景技术
布瓦西坦(brivaracetam,商品名Briviact),(2S)-2-[(4R)-2-氧代-4-丙基-1-吡咯烷基]丁酰胺,也称为UCB 34714,是由比利时UCB公司研制的一种新型抗癫痫药(AED),分别于2016年1月和2月被EMA和FDA批准上市,作为一款辅助治疗药物用于16岁及以上癫痫患者部分性癫痫发作的治疗。布瓦西坦属于第Ⅲ代新型抗癫痫药,是目前最畅销抗癫痫药物左乙拉西坦的结构衍生物。布瓦西坦是一种高选择性高亲和性的突出小泡蛋白2A配体,其抗癫痫作用是由多种机制产生的,主要作用机制是通过与中枢突触囊泡蛋白2A(synapticvesicle protein 2A,SV2A)结合影响突触功能,以及对电压依赖性钠离子通道的抑制作用所产生。因其具有很好的安全性和药动学性质,特别是中枢神经系统的良好耐受性,一直是癫痫治疗中的研究热点,拥有很大的市场应用前景。
目前布瓦西坦报道合成路线较多,主要包括通过手性色谱柱制备、手性拆分剂拆分以及不对称合成等方法。手性柱色谱制备方法文献报道较多,虞成功等对其合成方法进行了综述,这些方法需手性柱色谱拆分,分离制备成本高,生产规模有限,不能满足工业化生产的要求;王春燕等报道的手性拆分剂方法,选择了廉价易得的(R)-(+)-α-苯乙胺作拆分剂,de(非对映体过量百分率)值高,缺点是拆分后收率低,有超过50%的非对映异构体损失。刘九知等研究以不对称合成的方法来制备,但过程中使用柱层析纯化方法,不利于工业生产。综合以上几种化学合成方法可以看到化学合成工艺途径存在着收率低、成本高、步骤繁多、产物纯度较低、污染严重等问题。文献报道了利用蛋白酶拆分制备布瓦西坦中间体R-2-(2-叔丁氧基-2-氧代乙基)戊酸(Schule A,Merschaert A,Szczepaniak C,et al.ABiocatalytic Route to the Novel Antiepileptic Drug Brivaracetam[J].OrganicProcess Research&Development,2016,20(9)),生物催化具有反应温和,副产物少的优点。由于布瓦西坦专利将于2021年到期,目前市场需求量日渐加大,但目前布瓦西坦合成路线均存在一定的局限造成其目前原料药市场售价奇高,因此进行布瓦西坦合成工艺和有关物质研究,开发成本低廉、适合产业化放大工艺路线,制备高光学纯度布瓦西坦,具有极大的科研意义和经济价值。本发明利用新筛选的脂肪酶催化立体选择性水解反应拆分制备布瓦西坦手性中间体R-2-(2-叔丁氧基-2-氧代乙基)戊酸,催化剂易得成本低,ee值和转化率高,路线短操作简便。
(三)发明内容
本发明目的是为了解决现有合成方法的不足,通过筛选产脂肪酶微生物,提供一种廉价高效的立体选择性脂肪酶在生物法拆分RS-2-(2-叔丁氧基-2-氧代乙基)戊酸甲酯中的应用。
本发明采用的技术方案是:
本发明提供一种米曲霉脂肪酶,所述米曲霉脂肪酶氨基酸序列如SEQ ID NO.1所示,所述脂肪酶编码基因核苷酸序列为SEQ ID NO.2所示。
所述脂肪酶基因源自米曲霉WZ007(Aspergillus oryzaeWZ007),该菌保藏于中国典型培养物保藏中心,地址:中国武汉,武汉大学,430072,保藏编号:CCTCC No:M 206105,保藏日期:2006年10月8日,已在先前申请专利(中国专利CN 101186938A)中提交了相关菌种保藏信息并披露了其遗传资源来源。本发明从米曲霉WZ007出发,提取mRNA经反转录得到cDNA,根据NCBI查得的米曲霉基因组GenBank:XM_023233508.1碱基序列的基础上,设计上游引物为5'-CCATGGGCATGACAGCACACGAAGCCCTGA-3'和下游引物为5'-CTCGAGCTAATGTAACGCAACCCGAATATGC-3',进行PCR,得到碱基基因序列,测序结果为SEQ IDNO.2片段。
本发明还涉及含有米曲霉脂肪酶编码基因的重组载体,以及利用所述重组载体转化得到的重组基因工程菌,所述工程菌将SEQ ID NO.2所示基因连接至载体pET-28b(+),转化至大肠杆菌Rosetta感受态细胞,获得基因工程菌。
本发明提供一种所述米曲霉脂肪酶在拆分RS-2-(2-叔丁氧基-2-氧代乙基)戊酸甲酯制备布瓦西坦中间体R-2-(2-叔丁氧基-2-氧代乙基)戊酸中的应用(如图1所示,产物为布瓦西坦手性中间体),具体应用方法为:将含米曲霉脂肪酶基因的工程菌经发酵培养获得的湿菌体冷冻干燥后得到的脂肪酶粗酶粉为催化剂,以外消旋RS-2-(2-叔丁氧基-2-氧代乙基)戊酸甲酯为底物,以pH7.0磷酸缓冲溶液为反应介质,在20-60℃、1000rpm条件下进行拆分反应,反应完全后,将反应液分离纯化,制备获得布瓦西坦中间体R-2-(2-叔丁氧基-2-氧代乙基)戊酸。所述催化剂用量以缓冲液体积计为20g/L-100g/L(优选50g/L),所述底物终浓度为1%-3%(V/V)(优选1.75%)。
进一步,优选反应时间为5min-180min,更优选反应条件为45℃、1000rpm反应90min。
进一步,所述缓冲液为pH 7.0,0.2mM的Na2HPO4/NaH2PO4缓冲溶液。
进一步,所述催化剂按如下方法制备:将含米曲霉脂肪酶编码基因的工程菌(优选大肠杆菌Rosetta)接种在LB培养基中,37℃培养OD600至为0.4-0.6,加IPTG至终浓度0.02mM,30℃培养10-12h,菌液8000rpm,4℃离心10min,收集菌体,再用PBS缓冲液洗涤菌体2次,8000rpm,4℃离心10min,收集菌体,冻干(真空冷冻干燥机冷阱-80℃,真空度20Pa),得到脂肪酶粗酶粉;LB培养基组成:胰蛋白胨10g/L、酵母粉5g/L、NaCl 5g/L,溶剂为去离子水,pH值自然。
进一步,本发明所述反应液分离纯化的方法为:反应结束后,反应液先用2M NaOH水溶液调pH至9.0,加入等体积的环己烷萃取分离未反应的S-2-(2-叔丁氧基-2-氧代乙基)戊酸甲酯;然后用4M HCl水溶液将水相酸化至pH 2.0,加入等体积乙酸乙酯萃取,分液漏斗分离出有机相,有机相再经纯水洗涤两次,饱和NaCl洗涤两次,干燥,得到产物R-2-(2-叔丁氧基-2-氧代乙基)戊酸。
本发明脂肪酶氨基酸序列如SEQ ID NO.1所示:
MTAHEALNPIHPSVLPHLDPVFIKLYNENVANTPNKPIDLAILRSKYSVLYSYGTGPAPDPARIYDATVPGYNGDLIPVRVYEPSSPGPWPVHIDFHGGGMHSFPLLYPRIRRLMPTAGWGLGDLDTEAHICKHLSVKADVCVIDIGYRLVPEQPFPIGIQDSFAALEYIHAQGASKFNIDTTRISLGGVSAGGNIALIVAHLARDASIPLKLVAVGTPVIDDISKYASASESPYPSVQQMEHAPTLNWARLKWFDNLKWESLSSDVGLRKEQLDKISWYANAMNAPSFTNLPKTVIYTAGCDPLRDEGEAYAMKLVEGGNEVTLKRFEGVPHPFMHMDNDLWQAKEFIDKTAAHIRVALH。
由于氨基酸序列的特殊性,任何含有SEQ ID NO.1所示氨基酸序列的多肽的片段或其变体,如其保守性变体、生物活性片段或衍生物,只要该多肽的片段或多肽变体与前述氨基酸序列同源性在90%以上、且具有相同的酶活性,均属于本发明保护范围之列。具体的,所述改变可包括氨基酸序列中氨基酸的缺失、插入或替换;其中,对于变体的保守性改变,所替换的氨基酸具有与原氨基酸相似的结构或化学性质,如用亮氨酸替换异亮氨酸,变体也可具有非保守性改变,如用色氨酸替换甘氨酸。
本发明所述蛋白的片段、衍生物或类似物是指基本上保持本发明所述的蛋白酶相同的生物学功能或活性的蛋白,可以是下列情形:(Ⅰ)一个或多个氨基酸残基被保守或非保守氨基酸残基(优选的是保守氨基酸残基)取代,并且取代的氨基酸可以是也可以不是由遗传密码子编码的;(Ⅱ)一个或多个氨基酸残基上的某个基团被其它基团取代;(Ⅲ)成熟蛋白与另一种化合物(比如延长蛋白半衰期的化合物,例如聚乙二醇)融合;(Ⅳ)附加的氨基酸序列融合进成熟的蛋白而形成的蛋白序列(如用来纯化此蛋白的序列或蛋白原序列)。
所述蛋白可以是重组蛋白、天然蛋白或合成蛋白,可以是纯天然纯化的产物,或是化学合成的产物,或使用重组技术从原核或真核宿主(例如:细菌、酵母、高等植物、昆虫和哺乳动物细胞)中产生。根据重组生产方案所用的宿主,本发明的蛋白可以是糖基化的。本发明的蛋白还可以包括或不包括起始的甲硫氨酸残基。
本发明所述脂肪酶的编码基因核苷酸序列如SEQ ID NO.2所示:
ATGACAGCACACGAAGCCCTGAACCCTATCCACCCGTCCGTCCTGCCTCATTTGGACCCCGTCTTTATCAAACTCTACAATGAAAATGTCGCCAACACCCCCAACAAGCCCATAGACTTGGCCATTCTTCGATCAAAATATTCCGTGTTATATTCTTATGGTACCGGGCCAGCCCCCGATCCAGCTAGAATATACGATGCAACCGTGCCGGGATATAATGGCGATTTGATTCCAGTGCGAGTATACGAGCCATCGTCTCCGGGGCCTTGGCCGGTGCATATTGATTTTCATGGCGGTGGTATGCACTCCTTCCCTCTCTTGTATCCTCGGATCAGAAGACTAATGCCGACCGCAGGCTGGGGCCTTGGCGACCTCGACACTGAAGCTCATATCTGCAAGCATCTGTCCGTCAAAGCGGACGTTTGTGTAATCGACATTGGTTACCGGCTGGTCCCAGAACAGCCGTTTCCCATTGGCATCCAAGACTCCTTTGCTGCCCTGGAATACATTCATGCCCAGGGCGCTTCCAAGTTCAACATTGACACGACCCGCATCTCCCTTGGCGGTGTCTCAGCTGGAGGAAACATCGCCCTGATCGTGGCCCACCTTGCAAGGGATGCCAGCATCCCTCTGAAACTCGTCGCGGTGGGCACACCCGTCATTGACGATATCTCCAAGTACGCCTCTGCAAGCGAGTCTCCATACCCCTCTGTCCAACAGATGGAGCACGCGCCCACTCTCAACTGGGCCAGGTTGAAGTGGTTCGATAATCTCAAATGGGAAAGCCTTTCCAGCGATGTGGGTTTGAGGAAGGAGCAATTAGATAAAATCAGCTGGTATGCGAATGCAATGAATGCGCCTAGTTTCACCAACCTACCCAAGACGGTGATCTACACTGCTGGCTGTGATCCGCTACGAGATGAAGGAGAGGCGTACGCAATGAAGCTAGTGGAGGGTGGCAATGAAGTTACGCTTAAAAGGTTTGAAGGCGTGCCACACCCTTTCATGCATATGGATAACGACTTATGGCAGGCGAAGGAGTTTATAGACAAGACGGCTGCGCATATTCGGGTTGCGTTACATTAG。
由于核苷酸序列的特殊性,任何SEQ ID NO.2所示多核苷酸的变体,只要其与该多核苷酸具有70%以上同源性、且具有相同的功能,均属于本发明保护范围之列。所述多核苷酸的变体是指一种具有一个或多个核苷酸改变的多核苷酸序列。此多核苷酸的变体可以是天然发生的等位变异体或非天然发生的变异体,包括取代变异体、缺失变异体和插入变异体。如本领域所知的,等位变异体是一个多核苷酸的替换形式,它可能是一个或多个核苷酸的取代、缺失或插入,但不会从实质上改变其编码的氨基酸的功能。
另外,可与SEQ ID NO:2所示多核苷酸序列杂交的多核苷酸(至少具有50%同源性,优选具有70%同源性),也在本发明保护范围之列,特别是在严格条件下可与本发明所述核苷酸序列杂交的多核苷酸。所述“严格条件”是指:(1)在较低离子强度和较高温度下的杂交和洗脱,如0.2SSC,0.1%SDS,60℃;或(2)杂交时加用变性剂,如50%(v/v)甲酰胺,0.1%小牛血清,0.1%Ficoll,42℃;或(3)仅在两条序列之间的同源性至少在95%以上,更好是97%以上时才发生杂交。并且,可杂交的多核苷酸编码的蛋白与SEQ ID NO:1所示的蛋白有相同的生物学功能和活性。
与现有技术相比,本发明的有益效果主要体现在:本发明提供了一种米曲霉脂肪酶,该脂肪酶编码基因可与表达载体连接构建得到含该基因的胞内表达重组质粒,再转化至大肠杆菌菌株中,获得重组大肠杆菌,再利用重组大肠杆菌或重组脂肪酶为生物催化剂催化拆分外消旋2-(2-叔丁氧基-2-氧代乙基)戊酸甲酯,可以得到产物R-2-(2-叔丁氧基-2-氧代乙基)戊酸,对映体过量值>99%和转化率达到49.88%,产物R-2-(2-叔丁氧基-2-氧代乙基)戊酸的质量收率达到49.5%。目前现有的研究方法如手性色谱柱制备、手性拆分剂拆分以及不对称合成等的转化率为20%-30%,收率一般在10%-30%。本发明得到的转化率为49.88%,比现有方法高20%-30%;质量收率为49.5%,比现有方法高20%-40%。本发明生物催化的手性合成反应具有条件温和、效率高,高度的化学选择性、区域选择性以及对映体选择性等优点,而且生物催化过程具有无毒、无污染和能耗低等特点,是一种环境友好的合成方法。
(四)附图说明
图1脂肪酶对映选择性水解拆分RS-2-(2-叔丁氧基-2-氧代乙基)戊酸甲酯的反应示意图;
图2RS-2-(2-叔丁氧基-2-氧代乙基)戊酸甲酯的液相色谱图;
图3脂肪酶催化RS-2-(2-叔丁氧基-2-氧代乙基)戊酸甲酯水解反应90min的液相色谱图;
图4 R-2-(2-叔丁氧基-2-氧代乙基)戊酸经甲酯化反应的液相色谱图。
(五)具体实施方式
下面结合具体实施例对本发明进行进一步描述,但本发明的保护范围并不仅限于此,本领域的普通技术人员根据这些实施方式所做出的方法上的变换均包含在本发明的保护范围内。
实施例1米曲霉WZ007发酵,cDNA制备,脂肪酶基因克隆和工程菌构建
1、米曲霉WZ007发酵条件:斜面培养基(g/L):去皮马铃薯200,蔗糖20,琼脂20,溶剂为去离子水,pH自然。发酵培养基(g/L):蛋白胨20,橄榄油1%(V/V),KH2PO4 1,MgSO40.5,NaCl 0.5,溶剂为去离子水,pH 7.0。
将米曲霉(Aspergillusoryzae)CCTCC No:M 206105接种至斜面培养基,30℃培养6-8d,获得斜面菌种。米曲霉WZ007保藏于中国典型培养物保藏中心,地址:中国武汉,武汉大学,430072,保藏编号:CCTCC No:M 206105,保藏日期:2006年10月8日,已在专利申请CN101186938A中公开。
取斜面菌种接种至含100mL发酵培养基的500mL摇瓶中,在温度30℃,转速200rpm的恒温摇床培养12h,制成种子液。将1L种子液倒入含18L发酵培养基的30L发酵罐进行发酵培养,其中温度和转速分别控制在30℃和200rpm,发酵培养48h收集湿菌体。
2、米曲霉CCTCC No:M 206105cDNA制备:取1g上述米曲霉湿菌体,使用全式金公司的TransZol Up Plus RNA Kit试剂盒提取mRNA,再使用TOYOBO公司的ReverTra Ace qPCRRT Kit,高效率地合成适用于Realtime PCR的cDNA模板。
3、米曲霉脂肪酶基因的克隆:根据NCBI查得的米曲霉基因组
在GenBank:XM_023233508.1碱基序列的基础上,分别设计上游引物为5'-CCATGGGCATGACAGCACACGAAGCCCTGA-3'和下游引物为5'-CTCGAGCTAATGTAACGCAACCCGAATATGC-3',以cDNA为模板,进行PCR,得到核苷酸序列为SEQID NO.2所示的基因片段,测序验证正确。SEQ IDNO.2所示基因片段的编码蛋白氨基酸序列为SEQ ID NO.1所示。
4、大肠杆菌工程菌的构建:将该SEQ ID NO.2所示的基因片段连接至载体pET-28b(+),转化至大肠杆菌Rosetta感受态,获得含脂肪酶的大肠杆菌工程菌(记为工程菌M16)。
实施例2含脂肪酶的大肠杆菌工程菌(M16)的发酵培养
将实施例1获得的大肠杆菌工程菌(M16)接种在LB培养基中,37℃培养OD600至0.5(大概培养2h),加IPTG至终浓度0.02mM,30℃培养10-12h。300mL菌液8000rpm,4℃离心10min,收集菌体,再用PBS缓冲液洗涤菌体2次,8000rpm,10min,收集菌体。将收集的菌体使用的真空冷冻干燥机(冷阱-80℃,真空度20Pa)冻干后得到脂肪酶粗酶粉,记为脂肪酶M16,放于4℃冰箱保存。LB培养基组成:胰蛋白胨10g/L、酵母粉5g/L、NaCl 5g/L,溶剂为去离子水,pH自然。
实施例3酶催化拆分RS-2-(2-叔丁氧基-2-氧代乙基)戊酸甲酯的反应
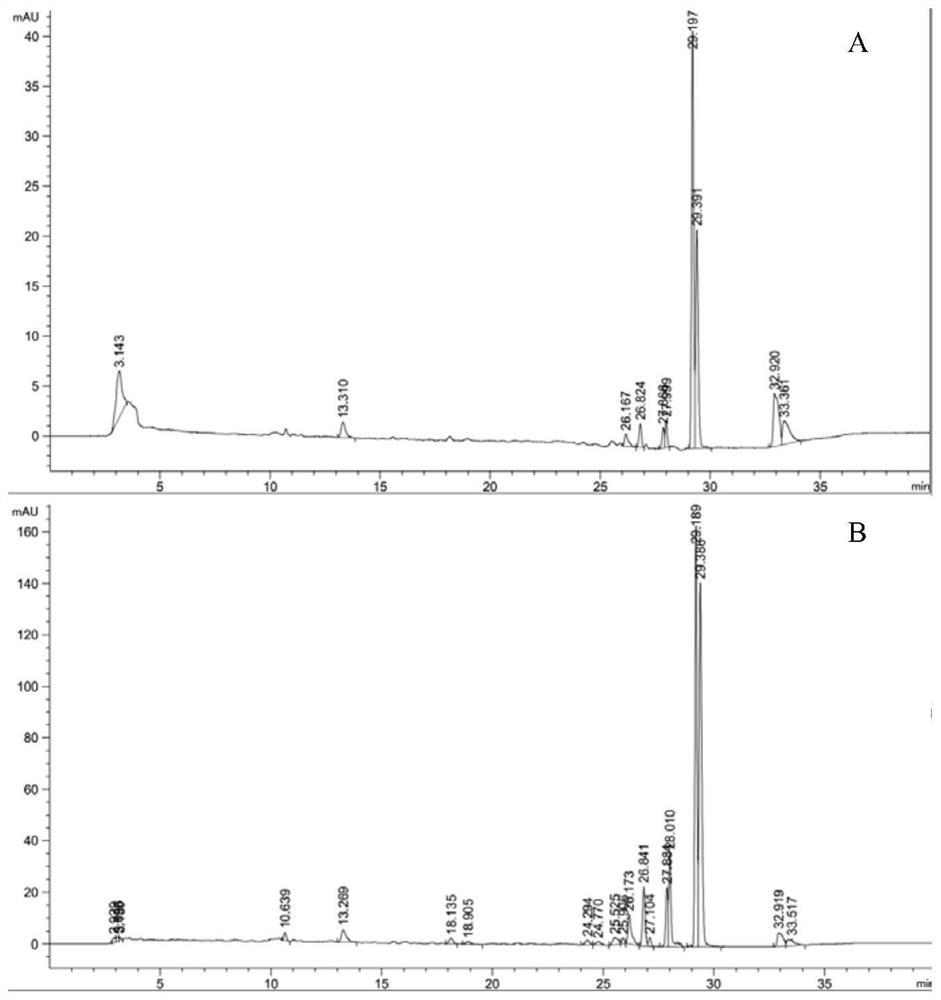
称取0.05g实施例2方法制备的脂肪酶M16于2mL EP管中,加入1mL PB(pH7.0、0.2mM)作为反应溶剂,然后加入17.5uL底物RS-2-(2-叔丁氧基-2-氧代乙基)戊酸甲酯,以不加脂肪酶为空白对照,置于45℃,1000rpm恒温混匀仪中反应90min。反应结束后,反应液先用2M NaOH水溶液调pH至9.0,加入等体积的环己烷,涡旋振荡器振荡充分萃取,离心(10000rpm,5min),得到含S-2-(2-叔丁氧基-2-氧代乙基)戊酸甲酯的有机相,取500uL经真空干燥,加入1mL流动相溶解,使用高效液相色谱检测菌体的立体选择性及酶催化水解活性。脂肪酶水解拆分反应,特异性水解了RS-2-(2-叔丁氧基-2-氧代乙基)戊酸甲酯得到所需的布瓦西坦中间体R-2-(2-叔丁氧基-2-氧代乙基)戊酸;不水解S-2-(2-叔丁氧基-2-氧代乙基)戊酸甲酯,所以在液相色谱图上只显示S-2-(2-叔丁氧基-2-氧代乙基)戊酸甲酯的峰。
具体液相分析条件:采用液相色谱仪Waters 1525型液相色谱仪;大赛璐CHIRALPAK AD-H(5μm,4.6×250mm)手性色谱柱,紫外光检测波长220nm,流动相正己烷:异丙醇=20:1(V/V);流速为0.5mL/min,柱温30℃,进样量10μL。S-2-(2-叔丁氧基-2-氧代乙基)戊酸甲酯标准品和R-2-(2-叔丁氧基-2-氧代乙基)戊酸甲酯标准品分别在9.87min和12.471min出峰(如图2所示)。酶水解拆分反应后,获得S-2-(2-叔丁氧基-2-氧代乙基)戊酸甲酯的液相色谱图(如图3所示),转化率49.68%,底物ee值>99%。
实施例4反应温度对酶动力学水解拆分RS-2-(2-叔丁氧基-2-氧代乙基)戊酸甲酯的影响
在pH 7.0,0.2mM的Na2HPO4/NaH2PO4缓冲溶液1mL中,加入终浓度50g/L的实施例2方法制备的脂肪酶M16,17.5μL RS-2-(2-叔丁氧基-2-氧代乙基)戊酸甲酯,于恒温混匀仪不同温度(20-60℃),1000rpm条件下反应90min,取反应液按实施例3方法检测S-2-(2-叔丁氧基-2-氧代乙基)戊酸甲酯的对映体过量值和转化率,结果见表1所示。
结果说明,在反应温度为45℃时,S-2-(2-叔丁氧基-2-氧代乙基)戊酸甲酯的对映体过量值最高,其ee值>99%。当反应温度高于45℃或者低于45℃时,催化转化率和对映体过量值都会下降,说明温度对脂肪酶M16的催化活性具有很大的影响。
表1反应温度对反应的影响
实施例5反应时间对酶动力学水解拆分RS-2-(2-叔丁氧基-2-氧代乙基)戊酸甲酯的影响
在pH 7.0,0.2mM的Na2HPO4/NaH2PO4缓冲溶液1mL中,加入终浓度50g/L实施例2方法制备的脂肪酶M16,17.5μLRS-2-(2-叔丁氧基-2-氧代乙基)戊酸甲酯,于恒温混匀仪45℃,1000rpm条件下反应不同时间(5min-180min),取反应液按实施例3方法检测R-2-(2-叔丁氧基-2-氧代乙基)戊酸甲酯的对映体过量值和转化率,结果见表2所示。
结果表明,反应90min后,产物S-2-(2-叔丁氧基-2-氧代乙基)戊酸甲酯的对映体过量值已达到最高,对映体过量值>99%,转化率为49.88%,当反应时间大于90min,转化率增大,将导致产物R-2-(2-叔丁氧基-2-氧代乙基)戊酸的对映体过量值降低。
表2反应时间对酶催化反应反应的影响
实施例6不同脂肪酶水解拆分RS-2-(2-叔丁氧基-2-氧代乙基)戊酸甲酯的效果比较
在pH 7.0,0.2mM的Na2HPO4/NaH2PO4缓冲溶液1mL中,加入终浓度50g/L的不同脂肪酶(实施例2方法制备的脂肪酶M16、Novozym 435、Lipozyme TL IM、Lipozyme RM IM),17.5μL RS-2-(2-叔丁氧基-2-氧代乙基)戊酸甲酯,于恒温混匀仪45℃,1000rpm条件下反应90min,取反应液按实施例3方法检测R-2-(2-叔丁氧基-2-氧代乙基)戊酸甲酯的对映体过量值和转化率,结果见表3所示。在45℃反应90min后,Novozym 435对RS-2-(2-叔丁氧基-2-氧代乙基)戊酸甲酯的两种构型能进行一定程度水解,但对底物两种构型没有对映体选择性,而LipozymeTL IM、LipozymeRM IM对底物没有水解活性和对映体选择性。当脂肪酶粗酶粉M16具有良好的立体选择性。
表3不同脂肪酶催化RS-2-(2-叔丁氧基-2-氧代乙基)戊酸甲酯的拆分效果
实施例7产物R-2-(2-叔丁氧基-2-氧代乙基)戊酸的分离提取及甲酯化
在50mL圆底烧瓶中加入10mL pH 7.0,0.2mM的Na2HPO4/NaH2PO4缓冲溶液,称取0.5g实施例2方法制备的脂肪酶M16,再加入终浓度1.75%(V/V)的RS-2-(2-叔丁氧基-2-氧代乙基)戊酸甲酯,用200mMNaOH水溶液进行流加滴定反应,控制反应pH维持在7.0,在磁力搅拌器45℃,600rpm条件下反应90min。反应结束后,反应液先用的2M NaOH水溶液调pH至9.0,加入等体积的环己烷萃取分离未反应的(S)-2-(2-叔丁氧基-2-氧代乙基)戊酸甲酯;然后用4M HCl水溶液酸化至pH 2.0,加入等体积乙酸乙酯萃取,分液漏斗分离出有机相,有机相再经纯水洗涤两次,饱和NaCl洗涤两次,干燥,得到最终产物,并称重0.071g,产物R-2-(2-叔丁氧基-2-氧代乙基)戊酸的质量收率达到49.5%。
R-2-(2-叔丁氧基-2-氧代乙基)戊酸的甲酯化:将所得R-2-(2-叔丁氧基-2-氧代乙基)戊酸,用1mL含10uL碘甲烷的DMF溶解,加入0.02g的K2CO3,在30℃、800rpm恒温混匀仪中反应3h,用500μL纯水洗涤后用1mL正己烷充分萃取离心取上相进行液相分析,液相色谱如图4所示,得出R-2-(2-叔丁氧基-2-氧代乙基)戊酸的对映体过量值>99%。
序列表
<110> 浙江工业大学
<120> 一种米曲霉脂肪酶及在制备布瓦西坦手性中间体中的应用
<160> 2
<170> SIPOSequenceListing 1.0
<210> 1
<211> 361
<212> PRT
<213> 米曲霉(Aspergillus oryzae)
<400> 1
Met Thr Ala His Glu Ala Leu Asn Pro Ile His Pro Ser Val Leu Pro
1 5 10 15
His Leu Asp Pro Val Phe Ile Lys Leu Tyr Asn Glu Asn Val Ala Asn
20 25 30
Thr Pro Asn Lys Pro Ile Asp Leu Ala Ile Leu Arg Ser Lys Tyr Ser
35 40 45
Val Leu Tyr Ser Tyr Gly Thr Gly Pro Ala Pro Asp Pro Ala Arg Ile
50 55 60
Tyr Asp Ala Thr Val Pro Gly Tyr Asn Gly Asp Leu Ile Pro Val Arg
65 70 75 80
Val Tyr Glu Pro Ser Ser Pro Gly Pro Trp Pro Val His Ile Asp Phe
85 90 95
His Gly Gly Gly Met His Ser Phe Pro Leu Leu Tyr Pro Arg Ile Arg
100 105 110
Arg Leu Met Pro Thr Ala Gly Trp Gly Leu Gly Asp Leu Asp Thr Glu
115 120 125
Ala His Ile Cys Lys His Leu Ser Val Lys Ala Asp Val Cys Val Ile
130 135 140
Asp Ile Gly Tyr Arg Leu Val Pro Glu Gln Pro Phe Pro Ile Gly Ile
145 150 155 160
Gln Asp Ser Phe Ala Ala Leu Glu Tyr Ile His Ala Gln Gly Ala Ser
165 170 175
Lys Phe Asn Ile Asp Thr Thr Arg Ile Ser Leu Gly Gly Val Ser Ala
180 185 190
Gly Gly Asn Ile Ala Leu Ile Val Ala His Leu Ala Arg Asp Ala Ser
195 200 205
Ile Pro Leu Lys Leu Val Ala Val Gly Thr Pro Val Ile Asp Asp Ile
210 215 220
Ser Lys Tyr Ala Ser Ala Ser Glu Ser Pro Tyr Pro Ser Val Gln Gln
225 230 235 240
Met Glu His Ala Pro Thr Leu Asn Trp Ala Arg Leu Lys Trp Phe Asp
245 250 255
Asn Leu Lys Trp Glu Ser Leu Ser Ser Asp Val Gly Leu Arg Lys Glu
260 265 270
Gln Leu Asp Lys Ile Ser Trp Tyr Ala Asn Ala Met Asn Ala Pro Ser
275 280 285
Phe Thr Asn Leu Pro Lys Thr Val Ile Tyr Thr Ala Gly Cys Asp Pro
290 295 300
Leu Arg Asp Glu Gly Glu Ala Tyr Ala Met Lys Leu Val Glu Gly Gly
305 310 315 320
Asn Glu Val Thr Leu Lys Arg Phe Glu Gly Val Pro His Pro Phe Met
325 330 335
His Met Asp Asn Asp Leu Trp Gln Ala Lys Glu Phe Ile Asp Lys Thr
340 345 350
Ala Ala His Ile Arg Val Ala Leu His
355 360
<210> 2
<211> 1086
<212> DNA
<213> 米曲霉(Aspergillus oryzae)
<400> 2
atgacagcac acgaagccct gaaccctatc cacccgtccg tcctgcctca tttggacccc 60
gtctttatca aactctacaa tgaaaatgtc gccaacaccc ccaacaagcc catagacttg 120
gccattcttc gatcaaaata ttccgtgtta tattcttatg gtaccgggcc agcccccgat 180
ccagctagaa tatacgatgc aaccgtgccg ggatataatg gcgatttgat tccagtgcga 240
gtatacgagc catcgtctcc ggggccttgg ccggtgcata ttgattttca tggcggtggt 300
atgcactcct tccctctctt gtatcctcgg atcagaagac taatgccgac cgcaggctgg 360
ggccttggcg acctcgacac tgaagctcat atctgcaagc atctgtccgt caaagcggac 420
gtttgtgtaa tcgacattgg ttaccggctg gtcccagaac agccgtttcc cattggcatc 480
caagactcct ttgctgccct ggaatacatt catgcccagg gcgcttccaa gttcaacatt 540
gacacgaccc gcatctccct tggcggtgtc tcagctggag gaaacatcgc cctgatcgtg 600
gcccaccttg caagggatgc cagcatccct ctgaaactcg tcgcggtggg cacacccgtc 660
attgacgata tctccaagta cgcctctgca agcgagtctc catacccctc tgtccaacag 720
atggagcacg cgcccactct caactgggcc aggttgaagt ggttcgataa tctcaaatgg 780
gaaagccttt ccagcgatgt gggtttgagg aaggagcaat tagataaaat cagctggtat 840
gcgaatgcaa tgaatgcgcc tagtttcacc aacctaccca agacggtgat ctacactgct 900
ggctgtgatc cgctacgaga tgaaggagag gcgtacgcaa tgaagctagt ggagggtggc 960
aatgaagtta cgcttaaaag gtttgaaggc gtgccacacc ctttcatgca tatggataac 1020
gacttatggc aggcgaagga gtttatagac aagacggctg cgcatattcg ggttgcgtta 1080
cattag 1086
一种米曲霉脂肪酶及在制备布瓦西坦手性中间体中的应用专利购买费用说明
![]()
Q:办理专利转让的流程及所需资料
A:专利权人变更需要办理著录项目变更手续,有代理机构的,变更手续应当由代理机构办理。
1:专利变更应当使用专利局统一制作的“著录项目变更申报书”提出。
2:按规定缴纳著录项目变更手续费。
3:同时提交相关证明文件原件。
4:专利权转移的,变更后的专利权人委托新专利代理机构的,应当提交变更后的全体专利申请人签字或者盖章的委托书。
Q:专利著录项目变更费用如何缴交
A:(1)直接到国家知识产权局受理大厅收费窗口缴纳,(2)通过代办处缴纳,(3)通过邮局或者银行汇款,更多缴纳方式
Q:专利转让变更,多久能出结果
A:著录项目变更请求书递交后,一般1-2个月左右就会收到通知,国家知识产权局会下达《转让手续合格通知书》。















动态评分
0.0