

IPC分类号 : C07K14/415I,C07K1/22I,C07K1/16I,C12N15/29I,C12N15/70I,C30B7/00I








专利摘要
本发明公开了水稻受体蛋白RGA5A_S的制备及结晶方法。蛋白RGA5A_S的制备方法为:利用原核表达系统表达水稻受体蛋白RGA5A_S,经过菌体裂解,裂解上清液依次过亲和层析柱、分子筛层析柱,最终得到纯化的目的蛋白;针对蛋白RGA5A_S包含金属离子结构域的特点,在重组菌诱导摇培时,向培养基中添加100μMZn2+,提高了蛋白稳定性。本发明还提供蛋白RGA5A_S的结晶方法。采用鸟枪法和九宫格法对蛋白结晶条件进行筛选优化,结果蛋白RGA5A_S在0.18‑0.22MNH4NO3和18%‑22%PEG3350条件下,所得晶体大、质量好、分辨率高,该方法重复性好。
权利要求
1.水稻受体蛋白RGA5A_S的制备方法,其特征在于,利用原核表达系统表达水稻受体蛋白RGA5A_S,经过菌体裂解,裂解上清液依次过亲和层析柱、分子筛层析柱,最终得到纯化的目的蛋白;
其中,重组菌诱导摇培时,培养基中添加100μM Zn
所述重组菌是将携带有水稻受体蛋白RGA5A_S编码基因的表达载体转化到大肠杆菌中构建得到的;
所述水稻受体蛋白RGA5A_S编码基因的序列如SEQ ID NO:1所示,所述表达载体为pETM13,所述大肠杆菌为Escherichia.coliBL21(DE3);
具体地,所述方法包括以下步骤:
(1)重组菌诱导表达:重组菌在37℃、220rpm摇培至菌液OD
(2)菌体裂解:收集经IPTG诱导的菌液,室温6000rpm离心8min,弃上清,收集菌体;用裂解缓冲液重悬菌体;超声裂解,开2s,关4s,功率400-600W,10min;2个循环;裂解物经12000-13 000rpm,4℃离心25-30min,得到细胞裂解上清液;
(3)亲和层析:将100mL细胞裂解上清液均分3份,分别加至3个经200mM ZnCl
(4)分子筛层析:分别收集60、500mM咪唑洗脱液,添加终浓度为4mM的DTT,分别浓缩至1mL;然后加样至
其中,步骤(2)中所用的裂解缓冲液为:20mM PBS,pH 7.0,500mM NaCl。
2.根据权利要求1所述的方法,其特征在于,步骤(3)所述洗脱缓冲液的基料为:20mMPBS,pH 7.0,500mM NaCl;咪唑浓度为0、20、60、500mM的洗脱缓冲液对应的洗脱体积为40、10、10、10mL。
3.根据权利要求1所述的方法,其特征在于,步骤(4)所用层析缓冲液的配方为:20mMTris-HCl,pH 8.0,150mM NaCl,5mM DTT和4mM EDTA。
4.根据权利要求1-3任一项所述的方法,其特征在于,步骤(2)~(4)在冰上或4℃条件下进行。
5.水稻受体蛋白RGA5A_S的结晶方法,其特征在于,用缓冲液将权利要求1-4任一项所述方法制备的目的蛋白配制成浓度6.4mg/mL的蛋白溶液,然后在0.2M NH
6.根据权利要求5所述的方法,其特征在于,晶种在0.18M NH
7.根据权利要求5或6所述的方法,其特征在于,所述缓冲液的配方为:20mM PBS,pH7.0,150mM NaCl,5mM DTT和4mM EDTA。
说明书
技术领域
本发明属于蛋白质结晶领域,具体地说,涉及水稻受体蛋白RGA5A_S的制备及结晶方法。
背景技术
水稻是世界三大经济作物之一,全球超过50%人口的主食(Liu and Wang,2016)。粮食安全成为当今全球经济生产头等大事,人口的持续增长是世界水稻产业发展的原始动力(周锡跃等,2010),至2030年世界水稻至少增长38%才能满足人类的需求(张璟等,2014)。因此,保障水稻稳定的产量与质量,直接关系粮食安全供给和人民温饱等问题,具有非常重要的战略意义。然而,稻谷的安全生产常年受病虫害的危害,其中,Magnaportheoryzae引起的稻瘟病是水稻病虫害中发生最为普遍、危害极大的真菌病害。稻瘟病菌的发生具有“点多面广”、发病面积大、发生区域集中、危害程度严重、区域间发病特点差异大等特点。不仅能够侵染水稻,还能侵染大麦、小麦、画眉草等多种禾本科作物和杂草,导致作物品质的下降。平均每年稻瘟菌引起水稻10%~30%的产量损失,而这足以解决2.12亿~7.42亿人口温饱问题(Fisher et al.,2012)。因此,稻瘟病菌被评为十大病原真菌之首(Dean et al.,2012)。目前,农业生产中主要采用抗病品种和化学杀菌剂对稻瘟病菌进行防控。随着人们生活水平和环保意识提高,越来越注重食物安全和品质,及其绿色环保的生活,化学农药的使用越来越受到限制。因此,选育抗病品种在控制稻瘟病的工作中变得至关重要;而选育抗病品种具有周期长、抗性稳定性差等缺陷。为了满足人们日益增长的粮食需求,迫切需要深入了解水稻受体与稻瘟病菌的协同进化分子机制,为抗病品种选育和品种布局提供科学、合理的理论基础。
蛋白质是构成细胞的基本有机物,也是生命活动的主要承担者和执行者,解析其分子结构对于了解其功能具有重要意义。结构生物学主要以生物学功能为主线,借助分子生物物理学和生物化学剖析生物大分子及其复合物三维结构,从原子的角度阐释蛋白质、核酸等发挥生物功能的分子机制。因此,深入了解蛋白质三维结构,不仅有利于人们深入了解蛋白质生物学功能,也为水稻抗病品种进行遗传改良和合理布局、药物设计、疫苗研发等应用领域提供理论基础。X-ray晶体学解析蛋白质三维结构是公认的最为可靠的方法。但是该技术一方面对蛋白质纯度(95%~99%)、稳定性、生物活性等有严格的要求;另一方面的瓶颈是结晶方法。因此,获得高质量的蛋白样品和稳定性好、可重复的高质量晶体对推进结构生物学解析蛋白质-蛋白质、蛋白质-核酸等生物大分子互作机制具有重要意义。
水稻和稻瘟病菌是植物与病原物相互作用的主要研究模式体系之一。目前已克隆水稻R基因25个和稻瘟病菌无毒基因11个,二者相互作用符合典型的“基因对基因假说”(Flor,1971)。已有研究表明,水稻中单个R基因不足以抵御病原菌的入侵,需要依赖成对的R基因共同参与防卫反应。水稻抗病基因Pia由两个R基因组成,RGA4和RGA5,翻译形成CC-NBS-LRR蛋白;在寄主体内可形成同源和异源复合物(Cesari et al.,2014;Cesari etal.,2013;de Guillen et al.,2015)。当RGA4单独存在的条件下,自发诱发寄主产生细胞过敏性坏死反应,该激活活性受到RGA5 的抑制。进一步分析发现,在RGA5蛋白C末端存在一个不保守的非LRR结构域(RGA5A_S结构域,包含金属离子结合结构域RATX1),直接参与识别两个序列不一致的效应因子AVR1-CO39、AVR-Pia。RGA5RATX1是参与病原菌识别的核心结合结构域,通过与AVR1-CO39、AVR-Pia直接结合,解除对RGA4的抑制,从而引发效应因子诱导的抗病反应(Cesari et al.,2014;Cesari et al.,2013)。RGA5还存在其他结构域直接参与效应因子AVR-Pia的识别,调节寄主免疫反应(Ortiz et al., 2017)。AVR1-CO39激发Pia水稻植株产生的活性氧爆发先天免疫反应,不仅能控制稻瘟病菌(M.oryzae)的入侵,还能够抑制革兰氏阳性细菌Xanthomonas oryzae pv. oryzae(Xoo)和Xanthomonas oryzaepv.oryzicola(Xoc)引起的水稻白叶枯病和水稻细菌性条斑病(Hutin et al.,2016)。目前,RGA5A_S识别稻瘟病菌效应因子 AVR1-CO39的互作机制尚不清楚,有待进一步研究。因此,需要针对水稻受体蛋白 Pia RGA5A_S提供一种晶体制备和生长技术,为深入了解植物受体R蛋白识别真菌效应因子提供结构基础。
前期研究发现,水稻受体蛋白RGA5A_S蛋白存在对温度敏感、易沉淀等问题。采用现有的蛋白质结晶方法,容易造成晶体生长机械手管道堵塞、蛋白质严重沉淀、晶体质量低等问题。
发明内容
本发明针对RGA5A_S蛋白对温度敏感,浓缩时易沉淀,结晶效率差以及晶体衍射质量低等问题,提供水稻受体蛋白RGA5A_S的制备及结晶方法。
为了实现本发明目的,本发明提供一种水稻受体蛋白RGA5A_S的制备方法,其是利用原核表达系统表达水稻受体蛋白RGA5A_S,经过菌体裂解,裂解上清液依次过亲和层析柱、分子筛层析柱,最终得到纯化的目的蛋白。其中,重组菌诱导摇培时培养基中添加有100μM Zn2+(ZnCl2)。
前述的方法,所述重组菌是将携带有水稻受体蛋白RGA5A_S编码基因的表达载体转化到大肠杆菌中构建得到的。
本发明蛋白RGA5A_S的氨基酸序列如SEQ ID NO:2所示。优选地,所述水稻受体蛋白RGA5A_S编码基因的序列如SEQ ID NO:1所示。
优选地,所述表达载体为pETM13。所述大肠杆菌为E.coli BL21(DE3)。
具体地,本发明水稻受体蛋白RGA5A_S的制备方法包括以下步骤:
(1)重组菌诱导表达:重组菌在37℃、220rpm摇培至菌液OD600值0.6~0.8,添加终浓度为100μM ZnCl2和0.1mM IPTG,18℃、180rpm诱导16~18h;
(2)菌体裂解:收集经IPTG诱导的菌液,室温6000rpm离心8min,弃上清,收集菌体;用裂解缓冲液(20mM PBS,pH 7.0,500mM NaCl)重悬菌体;超声裂解,开2s,关4s,功率400-600W,10min;2个循环;裂解物经12 000-13 000rpm,4℃离心25-30min,得到细胞裂解上清液;
(3)亲和层析:将100mL细胞裂解上清液均分3份,分别加至3个经200 mM ZnCl2预处理的亲和层析柱(chelating fast flow agarose column)上;穿出液重新挂柱一次;然后用含不同浓度咪唑0、20、60、500mM的洗脱缓冲液进行梯度洗脱;
(4)分子筛层析:分别收集60、500mM咪唑洗脱液,添加终浓度为4mM的 DTT(二硫苏糖醇),分别浓缩至1mL;然后加样至 75 10/300GL凝胶柱上,用层析缓冲液进行洗脱,合并收集的穿出液,穿出液中含有纯化的目的蛋白。
优选地,本发明所用亲和层析柱为chelating fast flow agarosecolumn。
步骤(3)所用洗脱缓冲液的基料为:20mM PBS,pH 7.0,500mM NaCl。
其中,咪唑浓度为0、20、60、500mM的洗脱缓冲液对应的洗脱体积为40、10、 10、10mL。
步骤(4)所用层析缓冲液的配方为:20mM Tris-HCl,pH 8.0,150mM NaCl,5mM DTT和4mM EDTA。
上述步骤(2)~(4)应尽量在冰上或4℃条件下进行。
本发明还提供水稻受体蛋白RGA5A_S的结晶方法,用缓冲液将上述方法制备的目的蛋白配制成浓度6.4mg/mL的蛋白溶液,然后在0.2M NH4NO3和20% PEG3350(v/v)存在的条件下,于16℃恒温培养箱中静置培养3-5天,获得微量晶体;然后以其为晶种,在0.18-0.22M NH4NO3和18%-22%PEG3350(v/v)存在的条件下,于16℃恒温培养箱中静置培养3-5天,得到水稻受体蛋白RGA5A_S 的单晶。
优选地,晶种在0.18M NH4NO3和20%PEG3350存在的条件下静置培养,获得单晶。
其中,所述缓冲液的配方为:20mM PBS,pH 7.0,150mM NaCl,5mM DTT 和4mMEDTA。
借由上述技术方案,本发明至少具有下列优点及有益效果:
(一)针对目的蛋白RGA5A_S包含金属离子结构域(具有MxCxxS基序,x 表示任意氨基酸)的特点,在重组菌摇培时向培养基中添加了Zn2+,对稳定蛋白的状态具有重要作用。
(二)采用鸟枪法和九宫格法对蛋白结晶条件进行筛选和优化,结果发现,蛋白RGA5A_S在0.18M NH4NO3和20%PEG3350条件下可获得大量单晶,所得晶体大、质量好、分辨率高,且该方法重复性好。
(三)由于蛋白RGA5A_S(不含有6×His标签),靠非保守的金属离子结合基序MxCxxS与Zn2+柱结合,结合力较6×His-tag差,因此,本发明选用含不同浓度咪唑的洗脱缓冲液对挂柱目的蛋白进行梯度洗脱,以保证低浓度下去除杂蛋白,高浓度下将高纯度目的蛋白完全洗脱。
附图说明
图1为本发明实施例2中获得的水稻受体蛋白RGA5A_S的微小晶体的偏光显微镜照片。
图2为本发明实施例2中获得的水稻受体蛋白RGA5A_S的单晶的偏光显微镜照片。
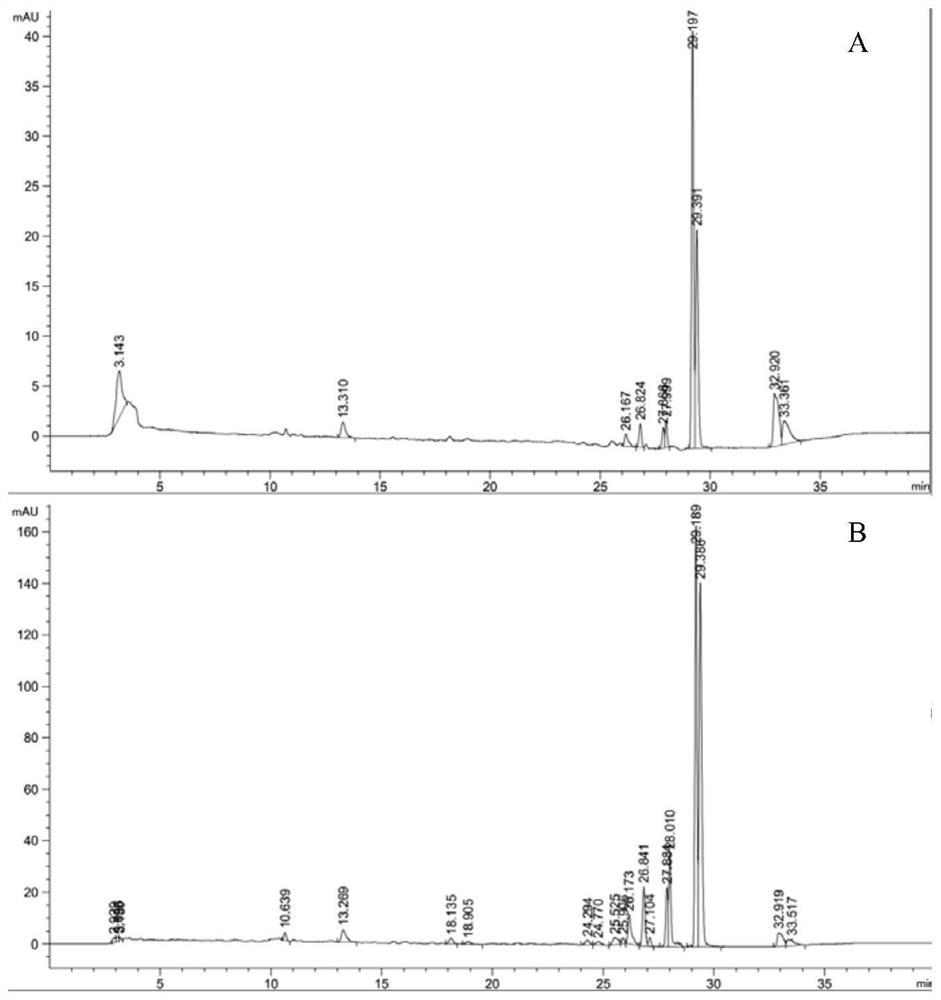
图3为本发明实施例2中水稻受体蛋白RGA5A_S晶体的的X-射线衍射数据。
具体实施方式
以下实施例用于说明本发明,但不用来限制本发明的范围。若未特别指明,实施例均按照常规实验条件,如Sambrook等分子克隆实验手册(Sambrook J&Russell DW,Molecular Cloning:a Laboratory Manual,2001),或按照制造厂商说明书建议的条件。
实施例1水稻受体蛋白RGA5A_S的制备
1、重组表达载体的构建
(1)引物设计
从NCBI上获得RGA5_RATX1(GenBank:BAK39930.1)目的序列,经密码子优化后合成基因序列。利用Sticky-end方法设计引物,根据选用载体的限制性酶切位点和目的序列限制性内切酶分析,选择限制性内切酶(NcoI/XhoI),利用DNAMAN 8.0设计两组引物。
F1-NcoI:catg CTGTCTAACA TGGAATCTGT
R1-XhoI:g CATAGTGCTG CACGGGTT
F2-NcoI:CTGTCTAACA TGGAATCTGT AGTAG
R2-XhoI:tcgag CATAGTGCTG CACGGGT
(2)PCR扩增目的片段和产物回收
采用Sticky-end的方法(Pham et al.,1998),将PCR扩增产物通过退火产生与载体末端互补的粘性末端。
(3)变复性-退火产生粘末端
利用NaNo Drop K550微量分光光度计检测PCR纯化产物浓度,按照摩尔比 1:1混合。利用PCR仪进行变复性,获得末端突出的双链DNA。
变性、复性条件:94℃变性5min,65℃变性15min,16℃冷却。
(4)酶切表达载体pETM13
对目的片段进行酶切,对载体进行双酶切,37℃孵育4-5h,
(5)载体与目的片段连接
在T4DNA连接酶的作用下,将变复性后产生粘性末端的PCR产物连接到酶切后具有相同末端的载体上,16℃孵育过夜。
(6)热击转化与抗性筛选
-80℃冰箱取出感受态细胞E.coli JM109/DH5α,至于冰上溶解;
向连接产物中加入50μL感受态细胞,冰上静置30min;
42℃水浴热击90s,立即于冰上静置2min;
超净台内加入500μL SOC培养基,37℃、220rpm振荡培养40min;
将培养基均匀涂至含有与载体相同抗性的LB平板,风干(超净台内操作);
倒置于37℃恒温培养箱过夜生长。
(7)重组子验证
随机挑取单克隆置于1mL相应抗性的LB液体培养基;
37℃、220rpm振荡培养5-10h;
菌液PCR验证插入片段是否正确;
扩增产物经琼脂糖凝胶检测后,选择条带与目的片段(SEQ ID NO:1)大小一致的菌液,送公司测序。利用DNAMAN8.0或NCBI将测序结果于目的序列进行比对,确认重组质粒中目的序列携带的酶切位点一致,碱基间有无移码和无点突变,则说明重组表达载体构建成功,可用于后续实验。
2、重组菌诱导表达及目的蛋白的分离纯化
(1)将上述重组表达载体转化到大肠杆菌E.coli BL21(DE3)中,在37℃、220 rpm摇培至菌液OD600值0.8,添加终浓度为100μM ZnCl2和0.1mM IPTG,18℃、 180rpm诱导18h。
(2)菌体裂解:收集经IPTG诱导的菌液,室温6000rpm离心8min,弃上清,收集菌体;用裂解缓冲液(20mM PBS,pH 7.0,500mM NaCl)重悬菌体;超声裂解,开2s,关4s,功率400-600W,10min;2个循环;裂解物经12 000-13 000rpm,4℃离心25-30min,得到细胞裂解上清液。
(3)亲和层析:将100mL细胞裂解上清液均分3份,分别加至3个经200 mM ZnCl2预处理的亲和层析柱上;穿出液重新挂柱一次;然后用含不同浓度咪唑0、 20、60、500mM的洗脱缓冲液进行梯度洗脱。
其中,所用亲和层析柱为chelating fast flow agarose column。
所用洗脱缓冲液的基料为:20mM PBS,pH 7.0,500mM NaCl。
咪唑浓度为0、20、60、500mM的洗脱缓冲液对应的洗脱体积为40、10、10、 10mL。
(4)分子筛层析:分别收集60、500mM咪唑洗脱液,添加终浓度为4mM的DTT,分别浓缩至1mL;然后加样至 75 10/300GL凝胶柱上,用层析缓冲液进行洗脱,穿出液中含有纯化的目的蛋白。
其中,所用层析缓冲液的配方为:20mM Tris-HCl,pH 8.0,150mM NaCl,5mM DTT和4mM EDTA。
实施例2水稻受体蛋白RGA5A_S的结晶方法
1、鸟枪法筛选蛋白质结晶条件
将实施例1制备的蛋白RGA5A_S溶解于缓冲液(20mM PBS,pH 7.0,150mM NaCl,5mMDTT,4mM EDTA)中,得浓度6.4mg/mL的蛋白溶液。然后进行鸟枪法晶体试剂盒筛选。16℃恒温培养箱中静置培养5天,在JCSG+试剂盒 (Hampton research)C4条件[蛋白溶液中添加有NH4NO3和PEG3350,终浓度分别为0.2M和20(v/v)%]可观察到微小晶体(图1)。而在其它晶体生长试剂条件下,几乎不能够获得晶体。
2、九宫格和接种策略优化晶体生长条件
为了获得高质量的衍射晶体,采用九宫格法优化NH4NO3和PEG3350浓度梯度,以步骤1中获得的微晶为晶种进行晶体生长条件优化。结果发现,RGA5A_S在 0.18M NH4NO3和20%PEG3350(v/v)存在条件下可获得大量单晶(图2)。在X- 射线下衍射,即可得到较高分辨率的衍射数据(一般为 左右的分辨率)(图3)。最后,利用相关的分析软件对收集衍射数据进行指标化(Index)、强度积分 (Integration)和归一化(Scaling)处理,即可得到该蛋白质的晶体结构。
虽然,上文中已经用一般性说明及具体实施方案对本发明作了详尽的描述,但在本发明基础上,可以对之做一些修改或改进,这对本领域技术人员而言是显而易见的。因此,在不偏离本发明精神的基础上所做的这些修改或改进,均属于本发明要求保护的范围。
参考文献
Cesari,S.,et al.(2014).″A novel conserved mechanism for plant NLRprotein pairs:the“integrated decoy”hypothesis.″Frontiers in plant science 5.
Cesari,S.,et al.(2013).″The rice resistance protein pair RGA4/RGA5recognizes the Magnaporthe oryzae effectors AVR-Pia and AVR1-CO39bydirect binding.″The Plant Cell 25(4):1463-1481.
de Guillen,K.,et al.(2015).″Structure analysis uncovers a highlydiverse but structurally conserved effector family in phytopathogenic fungi.″PLoS pathogens 11(10):e1005228.
Dean,R.,et al.(2012).″The Top 10fungal pathogens in molecular plantpathology.″Molecular plant pathology 13(4): 414-430.
Fisher,M.C.,et al.(2012).″Emerging fungal threats to animal,plant andecosystem health.″Nature 484(7393):186-194.
Flor,H.H.(1971).″Current status of the gene-for-gene concept.″Annualreview of phytopathology 9(1):275-296.
Hutin,M.,et al.(2016).″Ectopic activation of the rice NLR heteropairRGA4/RGA5confers resistance to bacterial blight and bacterial leaf streakdiseases.″The Plant Journal 88(1):43-55.
Liu,W.and G.-L.Wang(2016).″Plant innate immunity in rice:a defenseagainst pathogen infection.″National Science Review 3(3):295-308.
Ortiz,D.,et al.(2017).″Recognition of the Magnaporthe oryzae effectorAVR-Pia by the decoy domain of the rice NLR immune receptor RGA5.″The PlantCell Online 29(1):156-168.
张璟,et al.(2014).″2030年世界粮食发展及其对中美两国国家战略选择的影响.″世界农业(3):I0001-I0001.
周锡跃,et al.(2010).″世界水稻产业发展现状,趋势及对我国的启示.″農業現代化研究31(5):525-528。
序列表
<110> 中国农业大学
<120> 水稻受体蛋白 RGA5A_S 的制备及结晶方法
<130> KHP181112033.0
<160> 2
<170> SIPOSequenceListing 1.0
<210> 1
<211> 405
<212> DNA
<213> 水稻(Oryza sativa)
<400> 1
ctgtctaaca tggaatctgt agtagagtct gcgctgaccg gccagcgtac taagatcgtg 60
gttaaggtac acatgccgtg tggcaaaagc cgcgctaaag ctatggcgct ggcagcaagc 120
gtgaacggtg tggactctgt agaaattacc ggcgaagaca aagatcgcct ggtagttgta 180
ggccgtggca tcgatccggt tcgtctggtg gcgctgctgc gtgaaaaatg cggcctggcg 240
gaactgctga tggtagaact ggttgaaaaa gaaaagaccc agctggctgg cggcaaaaag 300
ggcgcgtaca agaaacaccc gacctacaac ctgagcccgt tcgattacgt tgaatacccg 360
ccgtccgctc cgattatgca ggacattaac ccgtgcagca ctatg 405
<210> 2
<211> 135
<212> PRT
<213> 水稻(Oryza sativa)
<400> 2
Leu Ser Asn Met Glu Ser Val Val Glu Ser Ala Leu Thr Gly Gln Arg
1 5 10 15
Thr Lys Ile Val Val Lys Val His Met Pro Cys Gly Lys Ser Arg Ala
20 25 30
Lys Ala Met Ala Leu Ala Ala Ser Val Asn Gly Val Asp Ser Val Glu
35 40 45
Ile Thr Gly Glu Asp Lys Asp Arg Leu Val Val Val Gly Arg Gly Ile
50 55 60
Asp Pro Val Arg Leu Val Ala Leu Leu Arg Glu Lys Cys Gly Leu Ala
65 70 75 80
Glu Leu Leu Met Val Glu Leu Val Glu Lys Glu Lys Thr Gln Leu Ala
85 90 95
Gly Gly Lys Lys Gly Ala Tyr Lys Lys His Pro Thr Tyr Asn Leu Ser
100 105 110
Pro Phe Asp Tyr Val Glu Tyr Pro Pro Ser Ala Pro Ile Met Gln Asp
115 120 125
Ile Asn Pro Cys Ser Thr Met
130 135
水稻受体蛋白RGA5A_S的制备及结晶方法专利购买费用说明
![]()
Q:办理专利转让的流程及所需资料
A:专利权人变更需要办理著录项目变更手续,有代理机构的,变更手续应当由代理机构办理。
1:专利变更应当使用专利局统一制作的“著录项目变更申报书”提出。
2:按规定缴纳著录项目变更手续费。
3:同时提交相关证明文件原件。
4:专利权转移的,变更后的专利权人委托新专利代理机构的,应当提交变更后的全体专利申请人签字或者盖章的委托书。
Q:专利著录项目变更费用如何缴交
A:(1)直接到国家知识产权局受理大厅收费窗口缴纳,(2)通过代办处缴纳,(3)通过邮局或者银行汇款,更多缴纳方式
Q:专利转让变更,多久能出结果
A:著录项目变更请求书递交后,一般1-2个月左右就会收到通知,国家知识产权局会下达《转让手续合格通知书》。














动态评分
0.0