

IPC分类号 : C12N9/02,C12N9/04,C12N9/16,C12N15/53,C12N15/55,C12P33/20,C12P33/00








专利摘要
本发明属于基因工程和生物合成领域,特别涉及甾体化合物的生物合成。本发明提供了在绿胶霉素类化合物合成中对侧链断裂具有关键作用的基因,分别命名为VidF、VidP和VidH及其编码多肽。所述多肽序列如SEQIDNO:1‑3所示;所述核苷酸序列如SEQIDNO:4‑6所示。
权利要求
1.一种分离的多肽,其包含:
(a)SEQ ID NO:1所示的多肽序列或者与其具有至少70%序列同一性,优选80%、85%、90%、93%、95%、97%、98%、99%序列同一性的多肽序列;或者
由SEQ ID NO:4所示的核酸序列或者与其具有至少70%序列同一性,优选具有80%、85%、90%、93%、95%、97%、98%、99%或100%序列同一性的核酸序列编码的多肽序列;和/或
(b)SEQ ID NO:2所示的多肽序列或者与其具有至少70%序列同一性,优选具有80%、85%、90%、93%、95%、97%、98%、99%序列同一性的多肽序列;或者
由SEQ ID NO:5所示的核酸序列或者与其具有至少70%序列同一性,优选具有80%、85%、90%、93%、95%、97%、98%、99%序列同一性的核酸序列编码的多肽序列;和/或
(c)SEQ ID NO:3所示的多肽序列或者与其具有至少70%序列同一性,优选具有80%、85%、90%、93%、95%、97%、98%、99%序列同一性的多肽序列;或者
由SEQ ID NO:6所示的核酸序列或者与其具有至少70%序列同一性,优选具有80%、85%、90%、93%、95%、97%、98%、99%序列同一性的核酸序列编码的多肽序列。
2.一种分离的多核苷酸,所述多核苷酸编码权利要求1所述的多肽,优选的,所述多核苷酸包含:
(i)SEQ ID NO:4所示的核苷酸序列或者与其具有至少70%序列同一性,优选具有80%、85%、90%、93%、95%、97%、98%、99%序列同一性的核酸序列;和/或
(ii)SEQ ID NO:5所示的核苷酸序列或者与其具有至少70%序列同一性,优选具有80%、85%、90%、93%、95%、97%、98%、99%序列同一性的核酸序列;和/或
(iii)SEQ ID NO:6所示的核苷酸序列或者与其具有至少70%序列同一性,优选具有80%、85%、90%、93%、95%、97%、98%、99%序列同一性的核酸序列。
3.表达盒,其包含根据权利要求2所述的多核苷酸。
4.载体,其包含权利要求2所述的多核苷酸或权利要求3所述的表达盒。
5.细胞,包含权利要求2所述的多核苷酸、权利要求3所述的表达盒或权利要求4所述的载体,所述细胞优选为真菌细胞,例如米曲霉细胞,更优选为多节孢属真菌,例如NodulisporiumSp.(65-12-7-1)。
6.一种绿胶霉素类化合物的合成方法,其包括将权利要求1所述的多肽或者权利要求1中至少两组所述多肽的组合与甾体化合物底物接触,优选的,所述甾体化合物底物在C17位连接有
7.一种催化甾体化合物C17-C20位键断裂的方法,其包括将权利要求1所述的多肽或者权利要求1中至少两组所述多肽的组合与甾体化合物接触,优选的,所述甾体化合物在C17位连接有
8.权利要求1所述的多肽、权利要求2所述的多核苷酸、权利要求3所述的表达盒、权利要求4所述的载体或权利要求5所述的细胞在绿胶霉素类化合物合成中的用途。
9.权利要求1所述的多肽、权利要求2所述的多核苷酸、权利要求3所述的表达盒、权利要求4所述的载体或权利要求5所述的细胞在催化甾体化合物C17-C20位键断裂中的用途。
10.一种试剂盒,其包含权利要求1所述的多肽、权利要求2所述的多核苷酸、权利要求3所述的表达盒、权利要求4所述的载体或权利要求5所述的细胞。
说明书
技术领域
本发明属于基因工程和生物合成领域,特别涉及甾体化合物的生物合成。
背景技术
自1945年绿胶霉素被发现以来(Brian,P.W.&McGowan.,J.C.Viridin:a highlyfungistatic substance produced by Trichoderma viride.Nature 156,144-145(1945)),大量研究表明这类化 合物具有光谱的抗真菌作用,抗炎作用以及抗细菌活性,特别地,还发现他们是纳摩尔级别 的PI3K基酶抑制剂,其中渥曼青霉素已经发展成为一种商业的PI3K抑制剂被广泛使用。由 于信号分子PI3K在肿瘤方面的重要作用,绿胶霉素类化合物作为PI3K抑制剂已被发现具有 重要的抗肿瘤活性,其中渥曼青霉素半合成类似物PX-886作为抗肿瘤药物最近已经进入II期 临床研究。由于这类化合物结构特殊,活性多样,已经引起人们的广泛关注。经过过去20年 期间不断开展的对呋喃甾体类化合物的全合成研究,已于2017年全合成了渥曼青霉素、左旋 绿胶霉素以及左旋绿毛菌醇。然而相比于化学合成,生物合成研究却一直未有实质性进展。
自上世纪60年代开始,人们便开始对绿胶霉素类化合物的生物合成机制展开研究。通过 喂养同位素标记的化合物mevalonate或者acetate,首次证明了绿胶霉素不是来自二萜途径, 而是源于甾体(Grove,J.F.Viridin.Part VI.Evidence for a steroidalpathway in the biogenesis of viridin from mevalonic acid.J.Chem.Soc.C.,549-551(1969);Hanson,J.R.&Wadsworth,H.J. Biosynthesis ofdemethoxyviridin.J.Chem.Soc.,Chem.Commun.,360-361.(1979);Simpson,T. J.,Lunnon,M.W.&MacMillan,J.Fungal products.Part 21.Biosynthesis of the fungalmetabolite,wortmannin,from[1,2-
发明内容
发明人经过多年的研究,分离获得了在绿胶霉素类化合物合成中对侧链断裂具有关键作 用的基因,分别命名为VidF、VidP和VidH。这三个基因能够有效的催化真菌体内绿胶霉素 类化合物的合成过程中甾体化合物的第17位碳与侧链20位碳之间之间连接键的断裂和/或修 饰,从而促进绿胶霉素类化合物合成的进行。这也是本领域首次分离出能够催化绿胶霉素类 化合物合成途径中C17-C20断裂的基因。
本发明的第一个方面提供了一种分离的多肽,其包含:
(a)SEQ ID NO:1所示的多肽序列或者与其具有至少70%序列同一性,优选80%、85%、 90%、93%、95%、97%、98%、99%序列同一性的多肽序列;或者
由SEQ ID NO:4所示的核酸序列或者与其具有至少70%序列同一性,优选具有80%、85%、 90%、93%、95%、97%、98%、99%或100%序列同一性的核酸序列编码的多肽序列;和/或
(b)SEQ ID NO:2所示的多肽序列或者与其具有至少70%序列同一性,优选具有80%、 85%、90%、93%、95%、97%、98%、99%序列同一性的多肽序列;或者
由SEQ ID NO:5所示的核酸序列或者与其具有至少70%序列同一性,优选具有80%、85%、 90%、93%、95%、97%、98%、99%序列同一性的核酸序列编码的多肽序列;和/或
(c)SEQ ID NO:3所示的多肽序列或者与其具有至少70%序列同一性,优选具有80%、 85%、90%、93%、95%、97%、98%、99%序列同一性的多肽序列;或者
由SEQ ID NO:6所示的核酸序列或者与其具有至少70%序列同一性,优选具有80%、85%、 90%、93%、95%、97%、98%、99%序列同一性的核酸序列编码的多肽序列。
本发明的第二方面提供了一种编码第一方面多肽的分离的多核苷酸,
在优选的实施方案中,所述多核苷酸包含:
(i)SEQ ID NO:4所示的核苷酸序列或者与其具有至少70%序列同一性,优选具有80%、 85%、90%、93%、95%、97%、98%、99%序列同一性的核酸序列;和/或
(ii)SEQ ID NO:5所示的核苷酸序列或者与其具有至少70%序列同一性,优选具有80%、 85%、90%、93%、95%、97%、98%、99%序列同一性的核酸序列;和/或
(iii)SEQ ID NO:6所示的核苷酸序列或者与其具有至少70%序列同一性,优选具有80%、 85%、90%、93%、95%、97%、98%、99%序列同一性的核酸序列。
本发明的第三个方面提供了一种表达盒,其包含根据本发明第二方面的多核苷酸。
本发明的第四个方面提供了一种载体,如表达载体,其包含了本发明第二方面所述的多 核苷酸或第三方面所述的表达盒。
本发明的第五个方面提供了一种细胞,包含本发明第二方面的多核苷酸、第三方面的表 达盒或第四方面的所述的载体。
在一个实施方案中,所述细胞为真菌细胞。
在一个优选的实施方案中,所述细胞为多节孢属真菌,例如Nodulisporium Sp.(65-12-7-1), 在另一个优选的实施方案中,所述细胞为米曲霉细胞
在一个实施方案中,所述细胞为细菌细胞,例如大肠杆菌细胞。
本发明的第六方面提供了一种绿胶霉素类化合物的合成方法,其包括将本发明第一方面 的任一组多肽、任选两组或者三组多肽的组合与甾体化合物底物接触。
在一个实施方案中,所述甾体化合物底物在C17位连接有 结构的侧链;
在一个优选实施方案中,所述甾体化合物底物是3-Dihydrovirone,其结构如下式所示:
其中优选的,所述甾体化合物底物是绿胶霉素类化合物的天然合成过程中的底物,更优 选的,所述底物是已经发生了C20-C22碳链断裂后的底物。
本发明的第七方面提供了一种催化甾体化合物C17-C20位键断裂的方法,其包括将本发 明第一方面的任一一组多肽、任选两组或者三组多肽的组合与甾体化合物接触,
在一个实施方案中,所述甾体化合物在C17位连接有 结构的侧链;
在一个具体实施方案中,所述甾体化合物是3-Dihydrovirone
在另一个具体实施方案中,所述甾体化合物为孕烷;
在另一个具体实施方案中,所述甾体化合物是孕酮。
在又一个实施方案中,所述甾体化合物是绿胶霉素类化合物的天然合成过程中的底物, 更优选的,所述化合物已经发生了C20-C22碳链断裂。
本发明的第八方面提供了本发明第一方面的多肽、第二方面的多核苷酸、第三方面的表 达盒、第四方面的载体或第五方面的细胞在绿胶霉素类化合物合成中的用途。
本发明的第九方面提供了本发明第一方面的多肽、第二方面的多核苷酸、第三方面的表 达盒、第四方面的载体或第五方面的细胞在催化甾体化合物C17-C20位键断裂中的用途。
本发明的第十方面提供了一种试剂盒,其包含本发明第一方面的多肽、第二方面的多核 苷酸、第三方面的表达盒、第四方面的载体或第五方面的细胞。
附图说明
图1a显示了渥曼青霉素(wortmannin),绿胶霉素(viridin)和去甲氧绿胶霉素(demethoxyviridin)的结构;图1b显示了本发明鉴定的来自Nodulisporium sp(65-12-7-1) 的去甲氧绿胶霉素合成基因簇。
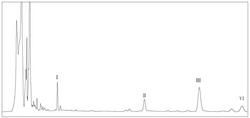
图2显示了vid基因缺失突变体的代谢物分析。图上部是来自vid基因缺失突变体的提取物的 HPLC图谱。图下部是从vid基因缺失突变体分离的化合物的结构。用蒸发光散射检测器 (ELSD)检测色谱图。
图3显示了本发明所发现的去甲氧绿胶霉素的生物合成过程中由VidF,VidP和VidH介导的 孕烷侧链切割机制。
图4显示了对纯化的重组VidF进行的SDS-PAGE分析结果,其中利用镍亲和层析纯化N-末 端使用6x His标记的VidF,重组VidF的分子量约为61kDa。
图5显示了酶法测定的HPLC分析。(a)同时添加化合物7、VidF和NADPH;(b)同时添加化合物7、灭活的VidF和NADPH;(c)仅添加化合物7和VidF;(d)化合物5(标准品); (e)化合物7和5的结构。
图6显示了喂食实验后进行HPLC分析的结果。其中,(a)携带vidP的米曲霉同时添加化合 物5;(b)携带空载体的米曲霉同时添加化合物5;(c)携带vidP的米曲霉;(d)携带vidH 的米曲霉同时添加化合物4;(e)携带空载体的米曲霉同时添加化合物4;(f)携带vidH的 米曲霉;(g)化合物5标准品;(h)化合物4标准品;(i)化合物9标准品;(j)化合物5, 4和9的结构。
图7显示了VidF,VidP和VidH介导对孕酮的侧链切割。(a)携带vidF+H+P的米曲霉(JA4) 同时添加化合物10;(b)携带空载体的米曲霉同时添加化合物10;(c)携带vidF+H+P的米曲霉;(d)化合物13标准品;(e)化合物12标准品;(f)化合物10标准品;(g)化 合物10-14的结构。
具体实施方式
还可进一步通过实施例来理解本发明,然而,要理解的是,这些实施例不限制本发明。 现在已知的或进一步开发的本发明的变化被认为落入本文中描述的和以下要求保护的本发明 范围之内。
术语“甾体化合物”,也被称为甾体,是一种存在于自然界中的一类天然化学成分,包括 植物甾醇、胆汁酸、C21甾类、昆虫变态激素、强心苷、甾体皂苷、甾体生物碱、蟾毒配基等。甾体化合物虽然种类众多,但是结构上具有相同的结构,均具有环戊烷多氢菲的基本骨架结构,同时在环戊烷多氢菲母核上通常带有两个角甲基(C-10、C-13)和一个含有不同的碳原子数的侧链或含氧基团如羟基、羰基等(C-17)(见下式)。
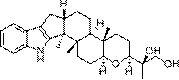
术语“绿胶霉素类化合物”,为由真菌产生的一系列C环芳香化且C-4和C-6位骈合一个 呋喃环的甾体化合物,是一类活性广泛且结构特殊的真菌次级代谢产物。这类化合物的结构 特点在于甾体骨架(A、B、C和D环)的C-4和C-6位间骈合一个环张力较大的呋喃环(E环)(如下式所示),因此这类化合物又称呋喃甾体类化合物(结构见下式)。
目前发现的天然来源的绿胶霉素类化合物均是真菌的次级代谢产物,曾在黏帚菌属(Gliocladium)、青霉属(Penicillium)、多节孢属(Nodulisporium)、木霉属(Trichoderma)、 膜盘菌属(Hymenoscyphus)、漆斑菌属(Myrothecium)和蓝状菌属(Talaromyces)中被分 离得到。按照其结构特点,可分为绿胶霉素(viridin)及其衍生物、渥曼青霉素(wortmannin) 及其衍生物以及viridin-related B-norsteroid类化合物。其中viridin及其衍生物的结构特点在 于结构中的甾体骨架不发生开裂和降解;wortmannin及其衍生物的结构特点在于结构中甾体 骨架的在C-2/C-3间发生开裂;viridin-relatedB-norsteroid类化合物的结构特点在于结构中的 甾体骨架的B环发生重排而形成一个五元环。
绿胶霉素化合物的生物合成前体是羊毛甾醇。而羊毛甾醇在C17位连接有包含七个碳的 侧链,因此在绿胶霉素化合物的合成过程中,需要伴随着对侧链剪切或者修饰。其中已知在 C20-C22位会发生一次侧链的断裂和修饰,因此推测形成具有 结构的侧链的甾体化合物 中间产物,由于作为终产物的绿胶霉素类化合物在第17碳直接仅直接与双键氧(=O)连接, 而不与碳原子连接,因此这就需要在最终形成绿胶霉素类化合物之前,在第17位碳发生断裂 和/或修饰。
在本发明所提供的VidF蛋白是一种单加氧酶(monooxygenase),能够在C17和C20之间 催化发生拜耳-维立格氧化重排反应(Baeyer-Villiger反应),其中拜耳-维立格氧化重排反 应(Baeyer-Villiger氧化重排反应)是在羰基和一个邻近烃基之间引入一个氧原子,得到相应 的酯的化学反应,经过上述反应,使得C17位侧链为 的甾体化合物在C17位转变为 结构。
在一个实施方案中,VidF蛋白所作用的底物是C17位侧链为 结构的甾体化合物。
在一个具体实施方案中,所述甾体化合物是指在绿胶霉素类化合物合成过程中的天然中 间产物,例如3-Dihydrovirone,并获得化合物5(Nodulisporiviridin N)
虽然VidF蛋白的天然功能是在合成绿胶霉素类化合物过程中,催化甾类化合物C17-C20 之间的修饰,但是VidF也同样可被用于对其他结构相近底物的修饰。例如,在一个实施方案 中,VidF蛋白的作用底物是孕烷或孕酮。
在一个具体实施方式中,VidF的多肽序列如SEQ ID NO:1所示,其编码核酸序列如SEQ ID NO:4所示。
本发明提供的VidP蛋白是一种脂酶,其能够使酯键水解,在本发明的一个实施方案中, VidP能够对经过VidF蛋白修饰后的C17侧链中的酯键进行切割,从而在C17位留下一个羟 基结构。
在一个具体实施方方案中,VidP以化合物5(即VidF的催化产物)为底物,产生在C17 位带有羟基的化合物4(Nodulisporiviridin M)。
在一个具体实施方案中,VidP的多肽序列如SEQ ID NO:2所示,其编码核酸序列如SEQ ID NO:5所示。
本发明提供的VidH是一种脱氢酶,在本发明的一个实施方案中,VidH能够对经过VidP 蛋白修饰后的C17位连接的羟基进行脱氢反应,从而在C17位形成“=O”结构。
在一个具体实施方案中,VipH以化合物4为底物,催化C17位的羟基转为=O,从而获得 化合物9(Nodulisporiviridin K)。
在一个具体实施方案中,VidH的多肽序列如SEQ ID NO:3所示,其编码核酸序列如SEQ ID NO:6所示。
在本发明的一个实施方案中,将VidF、VidP和VidH形成一个蛋白或基因的组合,同时 用于对甾体化合物C17位的修饰,在另一个实施方案中,可以选用两个蛋白或基因的组合, 例如VidF+VidP、VidP+VidH;在另一个实施方案中,可以仅适用一个蛋白或基因,例如仅适 用VidF、VidP或VidH。在不使用三种蛋白或基因的组合的情况下,需要针对组合的情况, 相应的调整甾体化合物底物的种类,而这是本领域技术人员在看到本公开后,容易自行调整 确定的。
实施例
材料与方法
1、对多节孢属真菌Nodulisporium Sp.(65-12-7-1)进行全基因组测序
多节孢属真菌Nodulisporium Sp.(65-12-7-1)(Zheng,Q.-C.etal.Nodulisporisteriods A and B,the first 3,4-seco-4-methyl-progesteroids fromNodulisporium sp.Steroids 78,896-901,2013; Zhao,Q.et al.NodulisporiviridinsA-H,bioactive viridins from Nodulisporium sp.J.Nat.Prod.78, 1221-1230,2015)保存在土豆琼脂PDA培养基上,接种到麦芽糖液体培养基中(3%maltose, 0.25%maltextract,0.15%yeast extract,0.2%KH2PO4,0.1%MgSO4,0.4%CaCO3),于28℃、转 速为180转/min的条件下发酵3天。过滤收集菌丝,液氮研磨,苯酚氯仿法提取DNA,使用Illumina HiSeq 2500system进行全基因组测序。测序数据利用软件SOAPdenovo version2.04 (http://soap.genomics.org.cn/soapdenovo.html)进行分析共得到381个重叠群(Contig),长度约 36.9Mb。使用软件AUGUSTUS(
2、构建基因敲除菌株
利用CRISPR/Cas9敲除系统对选定的三个基因(vidF,vidP,vidH)逐个进行敲除。具体方 法如下:
(1)体外转录纯化gRNA
以质粒pUCm-gRNAscaffold-eGFP(Zheng,Y.-M.et al.Development of aversatile and conventional technique for gene disruption in filamentous fungibased on CRISPR-Cas9technology. Sci.Rep.7,9250,2017)为模板,通过引物gRNA-vidF-F(SEQ ID NO:7)、gRNA-vidP-F(SEQ ID NO:8)、gRNA-vidH-F(SEQ ID NO:9)和eGFP-R(SEQID NO:10)扩增含有T7启 动子、20bp靶位点序列、gRNA骨架序列、tttt终止信号和eGFP的片段,加A后连接于T 载体,挑选阳性转化子时使用引物M13-F/eGFP-R,送阳性转化子测序验证,成功构建质粒 pUCm-gRNA-vidF,pUCm-gRNA-vidP和pUCm-gRNA-vidH。
随后,以上述质粒为模板,利用引物pUCm-F(SEQ ID NO:11)和gRNA-R(SEQ ID NO:12)进行PCR扩增并纯化,使用高保真的KOD Plus聚合酶扩增模板,经浓度测定和电泳检 测合格后,作为体外转录gRNA的DNA模板。然后进行体外转录。转录操作按照Promega 公司T7RiboMAX
然后使用Qiagen公司的RNeasy MinElute Cleanup Kit进行RNA纯化,具体过程如下: 操作过程中,使用“DNA and RNA away”将所需用到的器皿、移液枪、仪器等进行无酶处理, 使用DEPC处理过的枪头;在样品中加入80μL Rnase-free water,使体积为100μL;加入350 μL Buffer RLT混匀;加入250μL无水乙醇混匀;立即将溶液转移入吸附柱中,8000g离心 20s,倒掉收集管中溶液;将吸附柱放入新的2mL收集管中,加入500μL Buffer RPE,8000g,离心20s;加入500μL 80%乙醇,8000g,离心2min;将吸附柱放入一个新的收集管中 开盖,15000g,离心5min;将吸附柱放入新的1.5mL收集管中,在吸附柱中央加入20uL Rnase-free water,15000g,离心1min;紫外分光光度计测定浓度,-80℃保存。
(2)获得线性的neo抗性表达框
以质粒pBSKII-PtrPC-neo-TtrPC(Zheng,Y.-M.et al.Development of aversatile and conventional technique for gene disruption in filamentous fungibased on CRISPR-Cas9technology. Sci.Rep.7,9250,2017)为模板,使用引物PtrpC-XbaI-F(SEQ ID NO:13)/TtrpC-HindШ-R (SEQ ID NO:14),使用高保真的KOD Plus聚合酶进行扩增,获得抗性片段。
(3)制备原生质体,进行转染实验
1)将表达Cas9蛋白的多节孢属真菌Nodulisporium Sp.(65-12-7-1)的JN1001菌株(Zheng,Y. -M.et al.Development of a versatile and conventional techniquefor gene disruption in filamentous fungi based on CRISPR-Cas9technology.Sci.Rep.7,9250,2017)的孢子保存液 100μL加入到含有10mL DPY培养基的50mL试管中,30℃,150rpm,振荡培养1-2d。
2)将10ml的前培养液加入到装有100mL DPY培养基的100mL锥形瓶中,充分混合均匀, 30℃,120-150rpm,震荡培养24h。
3)制备10mL的TF solution 1,用0.22μm滤膜过滤50mL离心管。
4)取30mL左右的菌液,利用灭菌好的塞有棉花的注射筒过滤器进行过滤,将菌体压干, 用灭菌后的竹签将菌体取出,放入装有TF solution 1的离心管中。在30℃的恒温箱中振 荡破壁3h。
5)利用塞有棉花的注射筒过滤器,将破壁的菌液过滤到50mL的试管中。如果出现堵塞现 象,可以用竹签挑一下棉花表面,不要强用力,以免破坏原生质体。
6)加入等量的TF solution 2(约10mL),上下颠倒试管,轻轻混匀。常温,1500rpm离心 10min。倒去上清,加入5mL的TF solution 2,1500rpm离心10min。
7)用血球计数板在显微镜下计数原生质体的数目,利用TF solution 2调整原生质体浓度为 1-5×10
8)用1mL的枪头取200μL的原生质体溶液至15mL的离心管中,加入40-60μg gRNA与2-3μg neo抗性片段,轻轻地混匀。在冰上静置30min。向混悬液中分三次分别加入250μL,250μL,850μL的TF solution 3,每次加入后,都用1mL枪头轻轻吹吸。室温下静置20 min。
9)加入5mL的TF solution 2,上下颠倒试管,轻轻混匀。1500rpm离心10min,除去上清, 加入200μL的TF solution 2,用1mL的枪轻轻地混悬,下层培养基中。然后迅速加入5mL 50℃保温的上层培养基,快速混匀。
10)平板表面充分干燥后,用parafilm缠好,盖朝下进行培养3-7天。将获得转化株在不含 Sorbitol的筛选培养基上传代1-3次,获得稳定敲除菌株ΔvidF-JN1001,ΔvidP-JN1001和 ΔvidH-JN1001。
3、表达纯化VidF蛋白
1)首先构建重组质粒,以Nodulisporium sp.(No.65-12-7-1)cDNA为模板,使用引物 VidF-NdeI-F(SEQ ID NO:15)/VidF-NotI-R(SEQ ID NO:16)扩增vidF基因,然后将 片段连接到NdeI和NotI酶切的质粒pET-28a(+)(Novagen)中,构建质粒pET-28a (+)-vidF。
2)将重组质粒转化到宿主E.coli BL21-Codon Plus(DE3)(TaKaRa)中进行表达。
3)转化子接种至5mL含有50μg/mL Kanamycin的LB液体培养基中37℃,200rpm培养过 夜。
4)按1%接种量接入到含有50μg/mL Kanamycin的LB培养基中(500mL三角瓶装液量150 mL),37℃,200rpm培养3h左右至OD600值0.5-0.7,加入IPTG至终浓度为0.1mM, 18℃诱导表达16小时。
5)菌液4℃,5000g离心10min,弃掉上清液,然后用一定量的50mM Tris-HCl(8.0)清洗 菌体一次,再次离心后收集菌体,放置于-80℃冰箱中冻存。
6)取出冻存的菌体按40mL/1L菌液的比例添加裂解液(50mM Tris-HCl,200mMNaCl,5 mM imidazole,5%glycerol,pH 8.0)。充分混悬菌体,然后放置在冰上静置30min。
7)将菌液装在50mL离心管中,然后放置在冰水中。将超声破碎仪功率调至20%,工作5s, 休息5s,持续时间5分钟。
8)超声一次后,取出离心管上下颠倒几次后,继续超声破碎5min。
9)将破碎液经4℃,10000g离心10min,上层液经0.4μm过滤后用于进一步纯化。
10)取4mL Ni Sepharose
11)将过滤后的含有目标蛋白的上清液体添加至凝胶柱中,并收集流出液。
12)待液体流至表面时,用15-50倍柱体积的Wash Solution(50mM Tris-HCl,150mM NaCl,20 mM imidazole,5%glycerol,pH 8.0)洗脱杂蛋白。(收集后期Washsolution洗脱液,用于 电泳检测是否有目标蛋白洗脱出)
13)待液体流至表面时,用5mL的Elution Solution(50mM Tris-HCl,200mM NaCl,300mM imidazole,5%glycerol,pH 8.0)洗脱目标蛋白,并收集流出液。重复4次。
14)通过SDS-PAGE电泳检测目标蛋白主要富集于哪一份洗脱液中。
15)将含有目标蛋白的洗脱液转移至Amicon Ultra-4浓缩过滤管中,经4℃,4000g离心20 min,浓缩至1-2mL,然后向过滤管中继续添加透析液进行溶液交换,重复数次,直至稀 释比率约为1000。
16)将透析后的蛋白转移至离心管中,4℃,8000g离心3min,取上层蛋白液分装至离心管 中,先在液氮中速冻,然后放置在-80℃冰箱中保存。
17)浓度测定采用Bradford方法,牛血清蛋白为标准品。
4、体外酶活性实验
将纯化的VidF和底物3-dihydrovirone(7)在100μL溶液中反应,反应体系如下:50mM Tris-HCl(pH 8.0),125μM NADPH,0.25mM 7,1mM FAD,2μM VidF,25℃,150rpm震荡反应24h。反应液用乙酸乙酯终止,萃取两遍,提取物进行HPLC分析。
5、构建米曲霉A.oryzae NSAR1异源表达菌株
构建单基因表达菌株,分别从Nodulisporium sp.(No.65-12-7-1)基因组中扩增出目标基 因(VidF,VidP,VidH),通过In fusion连接方法连接到目标质粒pTAex3(Fujii,T.,Yamaoka,H., Gomi,K.,Kitamoto,K.&Kumaga,C.Cloning and nucleotide sequenceof the ribonuclease T1 gene(rntA)from Aspergillus oryzae and its expression inSaccharomyces cerevisiae and Aspergillus oryzae.Biosci.Biotechnol.Biochem.59,1869-1874,1995)上,形成重组质粒 pTAex3-vidF,pTAex3-vidP,pTAex3-vidH。
构建含有两基因(vidP,vidH)的表达质粒,分别从含有淀粉酶启动子和终止子的质粒 pTAex3-vidP,pTAex3-vidH中扩增出的DNA片段,然后用In fusion连接方法连接到pAdeA质 粒(Jin,F.J.,Maruyama,J.,Juvvadi,P.R.,Arioka,M.&Kitamoto,K.Developmentof a novel quadruple auxotrophic host transformation system by argB genedisruption using adeA gene and exploiting adenine auxotrophy in Aspergillusoryzae.FEMS Microbiol.Lett.239,79-85,2004)中, 构建两基因表达质粒pAdeA-vidP-vidH。
将单基因表达质粒pTAex3-vidP,pTAex3-vidH分别转染到A.oryzae NSAR1(Jin,F.J., Maruyama,J.,Juvvadi,P.R.,Arioka,M.&Kitamoto,K.Development of a novelquadruple auxotrophic host transformation system by argB gene disruptionusing adeA gene and exploiting adenine auxotrophy in Aspergillus oryzae.FEMSMicrobiol.Lett.239,79-85,2004)中,构建只 含VidP的米曲霉表达菌株JA1,和只含VidH的米曲霉表达菌株JA2将质粒pTAex3-vidF, pAdeA-vidP-vidH共同转染到A.oryzae NSAR1中,构建三基因(VidF,VidP,VidH)共表达 菌株JA3。
6、米曲霉喂养实验
将A.oryzae NSAR1转化菌株(JA1,JA2,或者JA3),接种到10mL DPY培养基中培养2天作为种子液,然后将种子液接种到50mL CD培养基中进行诱导表达,培养1天后,向培 养基中加入1mg底物(化合物5,4和9)(溶解在20μL DMSO中),继续培养4天后终止发 酵。发酵液用等体积的乙酸乙酯萃取2遍,提取物用HPLC进行分析。
7、化合物的分离纯化
化合物1和2的分离纯化:发酵2L的野生型菌株JN1001培养液用乙酸乙酯萃取2次。提取物经ODS中低压柱色谱洗脱(甲醇-水25:75,100:0,v/v),含有目标化合物的子馏分再经HPLC制备(25%乙腈-水,3mL/min)得到化合物1(18.2mg)和2(5.8mg)。
化合物3和4的分离纯化:发酵1L的敲除菌株ΔvidH-JN1001培养液用乙酸乙酯萃取2 次。提取物经ODS中低压柱色谱洗脱(甲醇-水25:75,100:0,v/v),含有目标化合物的子馏 分再经HPLC制备(25%乙腈-水,3mL/min)得到化合物3(5.5mg)和4(3.5mg)。
化合物5的分离纯化:发酵1L的敲除菌株ΔvidP-JN1001培养液用乙酸乙酯萃取2次。 提取物经ODS中低压柱色谱洗脱(甲醇-水35:65,100:0,v/v),含有目标化合物的子馏分再 经HPLC制备(45%甲醇-水,3mL/min)得到化合物5(9.5mg)。
化合物6,7和8的分离纯化:发酵3L的敲除菌株ΔvidF-JN1001培养液用乙酸乙酯萃取 2次。提取物经ODS中低压柱色谱洗脱(甲醇-水35:65,100:0,v/v),含有目标化合物6和 7的子馏分经HPLC制备(30%乙腈-水,3mL/min)得到化合物6(6.5mg)和7(9.5mg)。 含有目标化合物8的子馏分经HPLC制备(60%甲醇-水,3mL/min)得到化合物8(5.2mg)。
化合物11和14的分离纯化:发酵1L的米曲霉转化菌株JA3,喂养底物孕酮progesterone (150mg溶解在2mL DMSO),培养液用乙酸乙酯萃取2次。提取物经硅胶柱洗脱(氯仿- 甲醇100:0,80:20,v/v)得到3个馏分。馏分2(245.7mg)经中低压洗脱(甲醇-水5:5,8.5:1.5,10:0,v/v),含有目标化合物的子馏分再经HPLC制备(70%甲醇-水,3mL/min)得到化合物11(23.5mg)和14(14.5mg)
8、实施例涉及的部分化合物的结构、名称、编号以及核磁确认数据
化合物1核磁数据:
化合物2
化合物3
化合物4
化合物5
化合物6
化合物7核磁数据:.
化合物8核磁数据:.
化合物9
化合物11核磁数据:.
化合物14核磁数据:
实施例1候选基因的获得
通过能够富产去甲氧绿胶霉素(demethoxyviridin)的多节孢属真菌Nodulisporium Sp. (65-12-7-1)进行全基因组测序后,通过基因预测软件预测了相关的功能基因,并获得了与去 甲氧绿胶霉素合成相关的基因簇(图1b),发现了3个新基因VidF(多肽序列如SEQ ID NO: 1所示,核酸序列如SEQ ID NO:4所示)、VidP(多肽序列如SEQ IDNO:2所示,核酸序 列如SEQ ID NO:5所示)和VidH(多肽序列如SEQ ID NO:3所示,核酸序列如SEQ ID NO: 6所示)作为最终的候选研究基因。
实施例2基因敲除实验证明了这三个基因与C17位侧链的断裂和修饰密切相关
利用CRISPR/Cas9敲除系统对选定的三个基因(vidF,vidP,vidH)逐个进行敲除,在此前 构建的稳定表达Cas9蛋白的Nodulisporium sp.(No.65-12-7-1)JN001菌株(Zheng,Y.-M.et al. Development of a versatile and conventional technique forgene disruption in filamentous fungi based on CRISPR-Cas9technology.Sci.Rep.7,9250,2017)中构建得到了三个稳定敲除菌株 ΔvidF-JN1001,ΔvidP-JN1001和ΔvidH-JN1001。
从结果可以看出(图2),在对照组JN1001菌株中,高表达去甲氧绿胶霉素(化合物1), 但是在三个基因的敲除菌株中,均无法产生去甲氧绿胶霉素。其中在ΔvidF-JN1001中,富集 了化合物7或6,而在ΔvidP-JN1001中,仅能产生化合物5;而在ΔvidH-JN1001菌株中,则 富集产生了化合物3。比较三种敲除菌株各自富集的化合物(化合物7、5和3)可以看出三 者结构上的主要差别在于第17位碳侧链的结构,比较化合物7、5和3的结构,推测这三个 基因通过依次作用从而完成C17位侧链的断裂和修饰(图3)。
实施例3 VidF、VidP和VidH催化C17位侧链的断裂和修饰
为了确定所述三个基因的功能,在大肠杆菌中纯化获得重组的VidF(图4),进行体外酶 催化试验,结果表明VidF能够催化化合物7(3-Dihydrovirone)的C17-C20位置发生拜耳- 维立格氧化重排反应(Baeyer-Villiger反应),从而得到化合物5(Nodulisporiviridin N)(图5)。 由于VidP和VidH不能够在大肠杆菌中表达,于是分别将VidP和VidH导入米曲霉A.oryzae NSAR1中进行异源表达,通过底物喂养实验(分别喂养化合物5和4),证明了VidP能够以 化合物5(即VidF的催化产物)为底物,产生在C17位带有羟基的化合物4(Nodulisporiviridin M);而VipH则可以进一步以化合物4为底物,催化C17位的羟基转为=O,从而获得化合物 9(Nodulisporiviridin K)(图6),从而实现去甲氧绿胶霉素合成途径中C17位的修饰。
实施例4VidF、VidP和VidH的组合能够催化真菌内孕酮的侧链断裂
在真菌中孕酮侧链断裂的生物化学机制始终不清楚,接下来将三个基因(VidF,VidP, VidH)共同导入曲霉中,进行共表达,喂养孕酮(progesterone),结果发现孕酮C17位侧链 断裂,产生了不同的化合物(图7)。从而首次在基因水平上阐明了这种断裂机制。
序列表
<110> 暨南大学
<120> 甾体化合物侧链修饰基因及其应用
<130> 2018.4.1
<160> 16
<170> SIPOSequenceListing 1.0
<210> 1
<211> 541
<212> PRT
<213> Nodulisporium Sp. (65-12-7-1)
<400> 1
Met His Tyr Asp Leu Asp Ala Leu Ile Val Gly Ala Gly Phe Gly Gly
1 5 1015
Val Tyr Leu Leu His Gln Leu Arg Lys Glu Gly Leu Arg Ala Lys Ile
202530
Val Glu Ala Gly Asn Gly Leu Gly Gly Val Trp Asn Ser Asn Ser Tyr
354045
Pro Gly Ala Arg Ser Asp Thr Pro Ile Pro Thr Tyr Ser Leu Asn Ile
505560
Pro Glu Val Tyr Gln Thr Trp Thr Trp Asn Glu Arg Tyr Pro Gly Glu
65707580
Gln Glu Phe Lys Ala Tyr Phe Arg His Leu Asp Asn Val Leu Gly Met
859095
Ser Lys Asp Val Val Tyr Gly Glu Lys Val Ile Lys Ala Thr Phe Asp
100 105 110
Ala Ala Ile Asp Gly Trp Gln Ile Arg Thr Asp Arg Gly Thr Thr Phe
115 120 125
Thr Ala Arg Phe Phe Cys Pro Cys Leu Gly Phe Ala Ser Lys Asn Tyr
130 135 140
Thr Pro Asp Trp Pro Gly Ile Lys Asp Val Tyr Lys Gly Leu Ile Val
145 150 155 160
His Pro Asn Gln Trp Pro Ser Thr Gly Val Asp Met Lys Gly Lys Lys
165 170 175
Val Ala Val Val Gly Thr Gly Ala Thr Gly Val Gln Val Ala Gln Glu
180 185 190
Ala Ala Arg Asp Ser Ala Gln Leu Thr Cys Phe Val Arg Thr Pro Asn
195 200 205
Leu Ser Trp Pro Met Cys Gln Glu Lys Ile Asp Pro Glu Gln Tyr Glu
210 215 220
Lys Asp Arg Pro Leu Leu Glu Tyr Gln Leu Gly Thr Lys Arg Phe Thr
225 230 235 240
Thr Val Gly Gly Phe Leu Tyr Glu Glu Thr Glu Arg Val Leu Ala Asp
245 250 255
Asp Thr Pro Glu Glu Arg Glu Val Arg Leu Glu Glu Ala Tyr Lys Ile
260 265 270
Gly Gly Tyr Tyr Leu Phe Phe Cys Ala Tyr Ile Asn Val Leu Gly Asp
275 280 285
Gln Glu Gly Asn Asp Ala Leu Tyr Ala Phe Trp Arg Arg Lys Thr His
290 295 300
Glu Arg Met Thr Asp Lys Lys Lys Ala Glu Ile Leu Ala Pro Leu Lys
305 310 315 320
Pro Pro His Pro Phe Ala Gly Lys Arg Pro Ser Leu Glu Gln Asp Tyr
325 330 335
Tyr Glu Gln Met Asp Lys Pro His Val Thr Leu Val Asp Val Val Asn
340 345 350
Asn Pro Ile Thr His Leu Thr Ser Asn Ala Ile Val Thr Ala Asp Gly
355 360 365
Val Ala Tyr Glu Ala Asp Ile Ile Val Leu Ala Thr Gly Tyr Asp Ala
370 375 380
Ile Thr Gly Gly Leu Arg Asn Ile Glu Ile Thr Gly Leu Asp Gly Leu
385 390 395 400
Thr Leu Glu Glu Lys Trp Lys Asp Ser Thr Thr Ser Tyr Leu Gly Leu
405 410 415
Ala Thr Ser Gly Phe Pro Asn Met Phe Tyr Val Tyr Gly Pro Leu Ser
420 425 430
Pro Thr Ala Tyr Ala Ser Gly Pro Ala Val Val Glu Ser Gln Ala His
435 440 445
Trp Ile Met Asp Thr Ile Lys Lys Met Arg Asp Gln Asn Ala Thr Arg
450 455 460
Ile Asp Ala Thr Thr Lys Ala Glu Leu Glu Trp Arg Glu Lys Ile Glu
465 470 475 480
Thr Ile His Ala Tyr Thr Leu Arg Asp Lys Ile Glu Ser Ser Trp Tyr
485 490 495
Leu Gly Thr Asn Ile Pro Gly Lys Arg Arg Glu Val Leu Cys Tyr Ser
500 505 510
Gly Gly Leu Pro Met Tyr Arg Lys Glu Ile Phe Asp Ala Ile Lys Pro
515 520 525
Asp Trp Lys Gly Phe Thr Leu Lys Thr Thr Asp Ser Asn
530 535 540
<210> 2
<211> 354
<212> PRT
<213> Nodulisporium Sp. (65-12-7-1)
<400> 2
Met Asp Ala Ala Pro Arg His Tyr Ile Arg Gln Thr Phe Ala Ala Thr
1 5 1015
Arg Asn Leu Ile Ala Ile Phe Pro Ser Phe Ser Thr Asn Ile His Leu
202530
Ile Met Pro Leu Ala Tyr Asp Ser Glu Tyr Ala Lys Ala Leu Glu Pro
354045
Met Pro Ile Leu Ala Pro Arg Pro Thr Pro Lys Val His Asp Val Gly
Q:办理专利转让的流程及所需资料
A:专利权人变更需要办理著录项目变更手续,有代理机构的,变更手续应当由代理机构办理。
1:专利变更应当使用专利局统一制作的“著录项目变更申报书”提出。
2:按规定缴纳著录项目变更手续费。
3:同时提交相关证明文件原件。
4:专利权转移的,变更后的专利权人委托新专利代理机构的,应当提交变更后的全体专利申请人签字或者盖章的委托书。
Q:专利著录项目变更费用如何缴交
A:(1)直接到国家知识产权局受理大厅收费窗口缴纳,(2)通过代办处缴纳,(3)通过邮局或者银行汇款,更多缴纳方式
Q:专利转让变更,多久能出结果
A:著录项目变更请求书递交后,一般1-2个月左右就会收到通知,国家知识产权局会下达《转让手续合格通知书》。














动态评分
0.0